Parte 2: El origen de las adaptaciones de nuestro cuerpo a la marcha bípeda y el patrón de desarrollo. Una mirada desde la neurofisioterapia.
Nuestro cuerpo esta trufado en cada capítulo de explicaciones a cual más interesante de las adaptaciones que nuestra anatomía ha sufrido desde hace 7 millones de años. La mayoría estan relacionadas con las transiciones que nos llevaron desde una anatomía cuadrúpeda (probablemtente arborícora) a otra bípeda (1).
Neodarwinismo
La selección natural (SN) es el mecanismo que explica la mayoría de ellas, y la variación producida por las mutaciones al azar la fuente de la cual se nutre la selección natural para «escoger» los rasgos o caracteres que mejor se adaptaron a su nicho ecológico. Como insiste Arsuaga: «la naturaleza no busca pero encuentra». En otras palabras, no hay ninguna dirección en la evolución más allá de las presiones que un entorno genera para que unas características triunfen y sus portadores tengan mayor éxito reprodutivo.
Pero no todas las adptaciones son producto de la variación genética aleatoria ya que algunos rasgos pueden ser consecuencia de otros, es decir, una mutación que da lugar a una variación en una estructura (como la pelvis) tiene impacto en otras zonas del cuerpo que estan relacionadas con la posible zona mutada, y que incluso, puede interesar a todo el organismo (al tratarse de una unidad funcional). Algo similar a un efecto dominó.
En este sentido Arsuaga comenta en la pag.483: «Cada vez estoy más convencido de que las adaptaciones claves de los seres humanos cumplen múltiples funciones…descomponer el organismo en una constelación de rasgos físicos es una mala idea, porque un ser vivo es ante todo un sistema.»
Además de la selección natural, Arsuaga tambien menciona otras fuentes como la selección sexual que hace que determinadas caraterísticas sin valor adaptativo o funcional sean selecionadas por su atractivo para el otro sexo, como las plumas del pavo real, o la grasa acumulada en regiones concretas como el trasero.
Existen otras fuentes de variación al margen de la selección natural y la selección sexual?
Aunque la selección natural es la teoría dominante, la irrupción de la genética y otras disciplinas estan revolucionando el conocimiento de la evolución como dice Arsuaga en la pag 539, «…la genetica del desarrollo ha contribuido tanto a nuestro conocimiento de la evolución que ya no es posible separar la genetica de la embriologia, la paleontologia, clasifiación de las especies y la anatomia comparada, integrandose todas estas disciplinas en una nueva ciencia que se ha llamado Evo Devo (en ingles, evolucion y desarrollo)«.
A estas disciplinas hay que añadir la irrupción de la epigenética que esta abriendo un nuevo espectro de posibilidades para explicar desde otra perspectiva algunas adaptaciones relacionadas con estímulos procedentes del entorno y que pueden relacionarse con variaciones en los llamados genes reguladores (2).
El patron desarrollo: «lo que de verdad evoluciona»
Arsuaga apunta en la dirección que voy a desarrollar en este Post cuando en el capítulo sobre Las proporciones corporales, dice en la pag 186: «…No venimos al mundo con las proporciones adultas. Eso quiere decir que en nuestra especie se sigue un patrón de desarrollo diferente del de los chimpancés. O sea que se podría decir que lo que cambia en la evolución (lo que de verdad evoluciona) es el patron de desarrollo»
El patrón de desarrollo humano es la especialidad a la que he dedicado 28 años de profesión y a la que me sigo dedicando actualmente. Su estudio es esencial para detectar las desviaciones del patrón de desarrollo ideal (o típico para otros) que presentan los bebés, y realizar así un aborje precoz de estas posibles alteraciones. Aquí el tiempo es oro. En estos años he visto, y continuo viendo, incontables mutaciones que afectan al patrón de desarrollo, muchas con pronósticos vitales y funcionales catastróficos, y otras que permiten una calidad de vida aceptable con mayor o menor grado de dependencia.
El avance de la tecnología ha permitido ir identificando el origen de estas mutaciones, que se etiquetan genericamente como enfermedades raras, pero qué como el síndrome de Rett son cada vez son menos raras y más frecuentes. Nunca he conocido o tenido noticia de mutaciones al azar que afecten al patrón de desarrollo y que resulten ventajosas a sus portadores. Todas, sin excepción, son malas o muy malas.
El patrón de desarrollo y la marcha bípeda
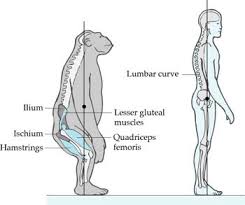
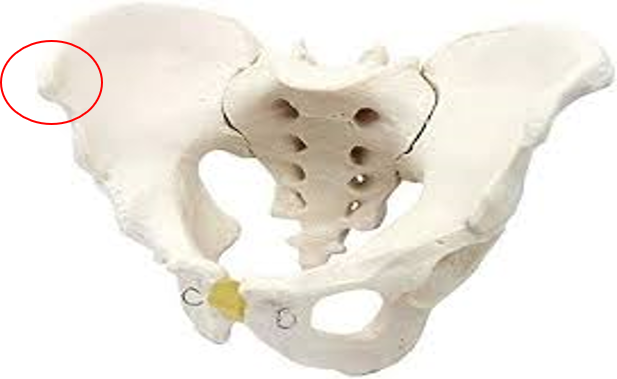
Hay consenso entre los paloantropologos que la primera mutación que cambió nuestra anatomía hacia la marcha bípeda se produjo en la pelvis. En concreto en la orientación de los iliacos, que pasaron de una disposición dorsal simiesca a una más lateral como en Lucy. Este hecho fue señalado por Washburn como el pistoletazo de salida hacia nuestra actual anatomía bípeda (3).
Pero en esta entrada no pretendo hablar de la estructura pélvica o hadware, sino de la función o software. En conreto de un movimietno nuevo que surgió en la pelvis a partir de ésta modificación estructural… o quizá fue al revés, ¿el nuevo movimiento podría haber sido el causasnte del cambio estrucutral? El eterno dilema entre estrucutra y función con el que tan familiarizados estamos los fisioterapeutas.
El nuevo movimiento que lo cambio todo: La retroversiónde la pelvis
Somos el único primate que posee este movimiento. En su obra Arsuaga lo define en el capítulo La pelvis y las nalgas en la pag 219, cuando menciona dos relieves anatómicos situados en la pelvis llamados Espinas Iliacas Antero Superiores (EIAS), dice: «la posición de la EIAS nos sirve para definir qué es anteversion y qué es retroversión de la cadera, términos que se utilizan habitualmente en la clase de pilates, en la de yoga y en muchas otras del gimnasio. En posición neutra las EIAS esta en el mismo plano frontal que el pubis. En anteversión, las EIAS se adelantan con respecto al pubis y en retroversión se retrasan.»
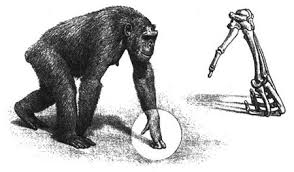
Estoy de acuerdo, pero solo es una parte de la definición. Si nos quedamos con esta parte los chimpancés, a partir de su «posición bípeda neutra» también tendrían retroversión porque pueden retrasar sus EIAS respecto al pubis aumentando la flexión de sus caderas y rodillas…y esto no es retroversión.
La retroversión es un movimiento complejo que implica tanto a la columna vertebral lumbar como a las caderas y ha sido explicado por el cirujano ortopédico francés Kapandji (4). Así, si tomamos una pelvis aislada, da igual como la coloquemos en el espacio que no hay forma de saber si esta en posición neutra, anteversión o retrovesión sin considerar la postura relativa que guarda con la columna y caderas.
Los chimpacés no tienen retroversión
En bipedestación, además de la definición de Arsuaga habría que añadir que para hacer retroversión la columna lumbar debe estar, bien en lordosis o axialmente extendida (no en cifosis), y las caderas en línea con el tronco.
Pero en nuestra evolución la implementación de la retroversión no se quedó ahí…su efecto fue como una enorme piedra lanzada al centro de un estanque, modificando toda la funcionalidad postural y motórica.
La retroversión pélvica en el patrón de desarrollo
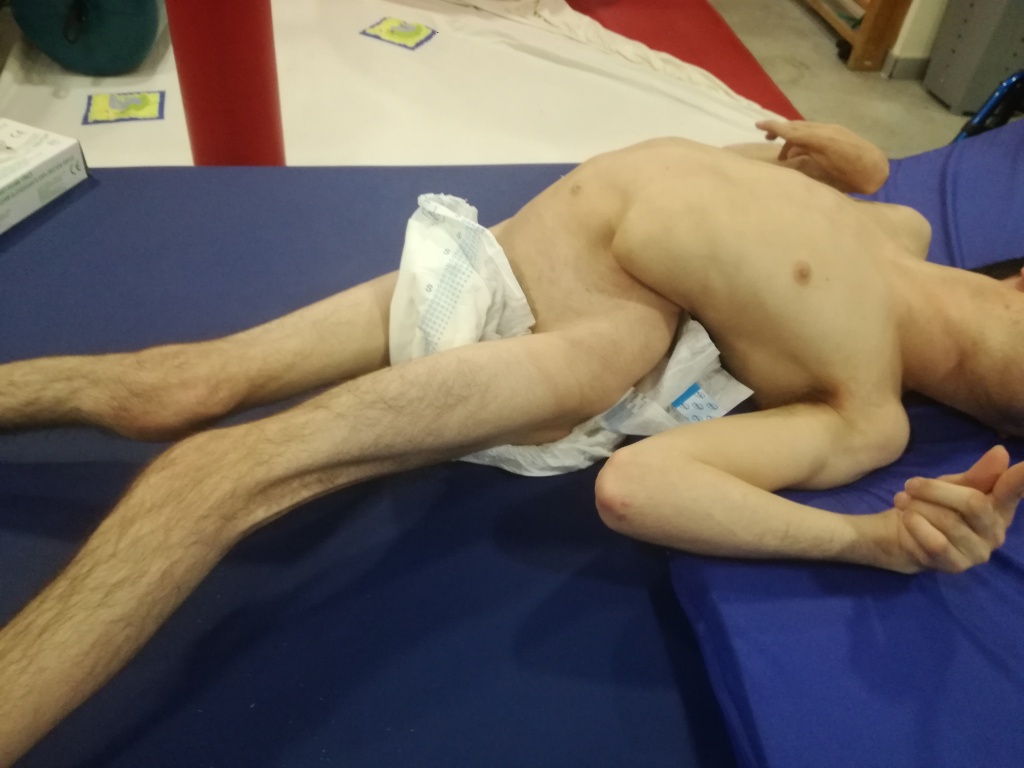
Aunque la marcha bípeda aparece en el entorno del año de vida, sus componentes estrucutrales son reconocibles en la etapa fetal, (no así en la embriológica), mientras que los cinemáticos (software) maduran más tarde. Los patrones esenciales que interesan a la marcha bípeda debutan a finales del primer trimestre, cuando, por primera vez, la columna vertebral (CV) se extiende axialmente de forma única entre los primates perdiendo así su cifosis prenatal (5).
Con esta extensión axial de la CV, a los 3 meses, la pelvis se posiciona en retroversión por primera vez, la retroversión que caracterizará nuestra marcha bípeda. De hecho, según un estudio de Gajewska y colegas (6) la posición de la pelvis a los 3 meses es un predictor fiable del proceso de desarrollo motor en nuestra especie.
A mediados del segundo trimestre surge otro patrón crítico para la marcha. Es el llamado patron de apoyo asimétrico sobre un codo, el cual supone un índice pronóstico clave para anticipar que el proceso va encaminado correctamente hacia la marha. ¿Por qué? porque la CV extendida axialmente rota por primera vez sobre su eje con la pelvis en retroversión. Elementos cinemáticos claves en la marcha bípeda de los que carece el chimpancé(5).
Tambien el debút del patrón cruzado que hacemos en la marcha, aparece mucho antes, en concreto a inicios del tercer trimestre con el patrón de la sedestación oblicúa, y toma forma de locomoción con el gateo dos meses más tarde. Sin embargo, habrá observado que algunos niños utilizan patrones alternativos al gateo para su locomoción, tanto en sustitución (como culear) como variantes.
Pero que un bebé se salte o configure un patrón de locomoción diferente al gateo típico no implica que carezca del patrón del gateo en su forma ideal, puesto que una vez que inicia la marcha, todas las alternativas al gateo y sus variantes son depuradas, es decir, desaparecen y no vuelven a repetirlas. Este curioso fenómeno se explica porque, tanto el gateo como la marcha, emplean los mismos núcleos de coordinacion neural (7).
Entonces, si el niño quiere volver utilizar estas variaciones pre-marcha como el «culeo» tendrá que hacerlo de un modo voluntario o consciente, es decir, lo autamático será la marcha, y si quiere desplazarse por el suelo de forma automática utilizará el gateo en su formato ideal. Esto sucede a cualquier edad y sin excepciones (patologias al margen).
Estos aspectos indican que todo nuestro proceso de maduración motriz durante el primer año de vida es un camino con una sola dirección: la marcha.
El patrón de desarrollo desde nuestra evolución.
El orden de aparición de los hitos del desarrollo en la ontogenesis desde el nacimiento hasta la marcha no siguen el orden de su aparición en la filogenesis (8). Es decir, primero sería la adopción temprana de la locomoción bípeda en los infantes presumiblemente por imitación, como postuló Richard Dawkins con el acuñamiento del concepto «meme» como la unidad de transmisión cultural que pasa de un cerebro a otro mediante el gesto, el comportamiento y en general cualquier otro fenómeno susceptible de ser imitado (9).
En este contexto, el comportamiento bípedo en una sociedad primate pre-bípeda (geneticamente hablando) sería un potente «meme» precursor de variación fenotipica susceptible de selección. Este «meme» supondría un ejercicio de rejuste postural en todos los componentes neurales y cinemáticos que ya estaban (y estan presentes) en la bipedia ejercida por todos los primates según un estudio de Druelle (10).
Aunque no sabemos la edad a la que los Australopitecos bebés alcanzaban la marcha bípeda, un estudio de Frémondière y colegas (11) realizando una simulación en 3d a partir de las medidas pélvicas de diferentes fosiles Austrolopitecus reveló que la estimación del peso de cerebro del bebé austrapopiteco sería de 110 gramos, el mismo que el de un bebé humano. Son 70 gramos menos de lo esperado en proporción al peso del cerebro de un australopiteco adulto.
Imagen del estudio de Takeshita donde compara reacciones posturales entre primates
Esto indica que «los recién nacidos de Australopithecus eran neurológicamente inmaduros y dependientes de la ayuda parental, de forma similar a los bebés humanos de hoy en día”, explica el autor. Por tanto, hay razón para pensar que su patron de desarrollo extrauterino hasta alcanzar la marcha bípeda fuese igualmente similar en proporción temporal a su periodo de gestación.
La tendencia al aumento de nuestra inmadurez se confirma en un estudio comparativo de madurez motriz entre los grandes simios (humanos incluidos) a través de la provocación de reacciones posturales automáticas, que muestran una distancia de un trimestre entre nosotros y los chimpancés para las mismas reacciones (12).
A la diferencia de casi dos trimestres para alcanzar la locomoción cuadrúpeda entre chimpancé y humano (gateo) habría que sumar un trimestre más al humano, qué es lo que tardamos en alcanzar la marcha bípeda. Con estos datos se podría estimar que el bebé australopiteco alcanzaría su marcha específica (la bípeda) más tarde que el chimpancé y antes que nosotros, esto es, entre el tercer y cuarto trimestre. Un período que se iría retrasando con el incremento del cerebro, naciendo, por consiguiente, progresivamete más inmaduro.
Reordenación de los hitos motores
Representación de Homo Habilis trabanado la piedra
Así, componentes específicos de la marcha bípeda moderna como la retroversión de la pelvis y la extensión axial surgirian en los primeros infantes pre-bípedos con la propia adopción de la marcha, no antes. Estos nuevos componentes cinemáticos, en el nuevo plan de desarrollo, los encontramos como ya se ha dicho, reubicados a finales del primer trimestre.
También, los componentes cinemáticos adquiridos en la filogenia después de la instauración de la marcha bípeda en el genotipo como es el agarre radial, de inicio presumible en Homo Habilis, (al ser un componente necesario para la confección de lascas) son igualmente reubicados en el plan de desarrollo.
En este caso, se calcula una diferencia de más de un millon de años después de adquirir la marcha, y resulta, que en el plan de desarrollo actual el agarre radial aparece al final del 2º trimestre, mientras que la marcha lo hace en el 5º. En otras palabras, la ontogenesis no solo resume la filogenesis, sino que tambien la reorganiza.
Siguiendo una metáfora informática sería como cuando el sistema operativo de tu ordenador o móvil necesita una actualización de software. Esta se instala sobre la anterior versión con el sistema en funcionamiento. Posteriormente, para que la actualización se ejecute plenamente es necesario reiniciar el ordenador para que las nuevas especificaciones se insertenen en el «lugar» asignado y con las interacciones que hagan funcionar de forma armónica y coherente todo el sistema ya actualizado.
Vamos, que las actualizaciones no se ubican al final de la «cola» por orden de llegada. Del mismo modo, tampoco lo hacen las actulizaciones que sufre el genotipo, sino que su ejecución esta mediada por modificaciones en los genes reguladores sin tener que seguir un orden secuancial según su entrada cronológica.
Marcadores epigenéticos y marcadores anatómicos
Lo fascinante de este proceso es que una vez que fuese asimilado el comportamiento bípedo en las crías a causa de la presión ambiental ejercida por una comunidad bípeda de uso y costumbre según Dawkins, el plástico cerebro y cuerpo de los pequeños iría haciendo cada vez más eficiente el patron neural de marcha bípeda mediante variaciones miofasciales y osteomusculares. Un trabajo de Hirasaki y colegas ha demostrado que esta plasticidad también funciona a la inversa entrenando macacos en la marcha bípeda desde su nacimiento (13).
De este modo, marcadores epigenéticos generados por el patron neural, y estrucutras anatómicas modificadas en el proceso bípedo (espinas, tubérculos, abulsiones, torsiones ect) quedarían asociadas en el genotipo. En consecuencia, para las siguientes generaciones se reorganizaría el patron de desarrollo ya en la etapa embrionaria para reubicar todos los patrones al servicio de la nueva locomoción asimilada (14).
Parece Lamarkismo…pero es herencia epigenética transgeneracional
Este relato suena sospechosamente a la herencia de los caracteres adquiridos de Lamark pero hay otra explicación acorde con la Síntesis Evolutiva Extendida, que integra la herencia epigenética transgeneracional, y que actúa esencialmente en el periodo fetal. Periodo que abarca el proceso ontogenético motriz que sigue al nacimiento inmaduro hasta la adquisición de la marcha desde, al menos, australopitecus. De hecho, como definió el zoologo suizo Adolf Portmann, en el primer año de vida somos literalmente «fetos extrauterinos» (15), y por consiguiente, altamente sensibles a la presión de nacer en un entorno «viralmente» bípedo.
En consecuencia, no sería estrictamente el cambio fenotípico adaptativo a la bipedia lo que se transcribiría al genotipo, sino el patron de metilación que hace que se exprese ese cambio.
El asombroso descubrimiento de los marcadores anatómicos vinculados a los marcadores epigenéticos
Entre los años 50 y 70 del pasado siglo un neuropediatra checo llamado Vaclav Vojta descubrió una forma de activar estos marcadores epigenéticos (que obviamente desconocía) provocando la expresión del programa neural de la marcha bípeda con todos los contenidos cinemáticos que pueden observarse durante la ontogenesis postural hasta consecución de la marcha(16). Curiosamente el acceso a estos marcadores fue a través de la presión de zonas concretas del cuerpo (marcadores anatómicos) con ejes vectoriales definidos (17).
Vojta descubrió díez de estas zonas, y desarrolló una metodología que actualmente utilizamos los fisoterapeutas de todo el mundo para evocar patrones específicos de la marcha humana, tanto en bebés como en adultos. Desde su descubrimiento, sus fundamentos neurofisiológicos han sido demostrados en estudios científicos recientes del máximo nivel utilizando la tecnología más avanzada (18-23). Sin embargo, los estrechos vínculos que estos trabajos guardan con nuestra evolución aún se desconocen para la comunidad científica que estudia nuestros orígenes.
Consideraciones finales
Postulamos la existencia de marcadores epigenéticos que registraron en el genotipo los patrones neurales específicos que configuran el patrón de marcha bípeda, almacenando, en el mismo «pack» las zonas anatómicas que más intensamente fueron modificadas hacia el morfotipo bípedo. Esta hipótesis no modifica en esencia los postulados actuales sobre la adquisición del bipedismo que impregna la obra de Arsuaga, tan solo busca incorporar una mirada desde un campo inédito para las ciencias que estudian la evolución como es el de la Neurorehabilitación.
Percibir el efecto en nuestro cuerpo de la actvación de estos marcadores epigenéticos es posible para cualquier persona, (de la mano un fisioterapeuta especializado en Vojta), y es, probablemente, la forma más eficaz e intensa de experimentar nuestra evolución hacia el bipedismo en nuestro propio cuerpo. Como dice Arsuaga: «todos tenemos un cuerpo» y en él esta escrita toda nuestra historia evolutiva al alcance de quien quiera en leerla.
Bibliografia
Juan Luis Arsuaga. Nuestro Cuerpo. Ed Destino 2023.
2. Mulligan CJ. Insights from epigenetic studies on human health and evolution. Curr Opin Genet Dev. 2018 Dec;53:36-42. doi: 10.1016/j.gde.2018.06.008. Epub 2018 Jul 4. PMID: 30015224.
3. Washburn SL. 1968. The study of human evolution. Eugene, OR: Oregon State System of Higher Education.
5. Vojta V, Schweizer E. El descubrimiento de la motricidad ideal. 2011. Ediciones Morata.Vojta.
6. Gajewska E, Sobieska M, Moczko J. The position of the pelvis in the 3rd month of life predicts further motor development. Hum Mov Sci. (2018) 59:37–45. doi: 10.1016/j. humov.2018.03.007
7. Dietz V. Quadrupedal coordination of bipedal gait: implications for movement disorders. J Neurol. 2011 Aug;258(8):1406-12.
8. Gould Jay Stephen. Ontogenia y filogenia. La ley fundamental biogenetica. Editorial crítica 2010.
10. Druelle F, Aerts P, Berillon G. The origin of bipedality as the result of a developmental by-product: The case study of the olive baboon (Papio anubis). J Hum Evol. 2017;113:155-161. doi:10.1016/j.jhevol.2017.07.010
11. Frémondière, P., Thollon, L., Marchal, F. et al. Dynamic finite-element simulations reveal early origin of complex human birth pattern. Commun Biol5, 377 (2022). https://doi.org/10.1038/s42003-022-03321-z.
12. Takeshita H, Tanaka, M, Matsuzawa T. Development of Postural Reactions in Infant Japanese Monkeys. The Annual of animal Psichology, 33 (1984)
13. Hirasaki E, Ogihara N, Hamada Y, Kumakura H, Nakatsukasa M. Do highly trained monkeys walk like humans? A kinematic study of bipedal locomotion in bipedally trained Japanese macaques. J Hum Evol. 2004 Jun;46(6):739-50. doi: 10.1016/j.jhevol.2004.04.004. PMID: 15183673.
14. Alice Roberts. La increíble probabilidad del ser. Pasado & Presente (2018).
15. Markus Ritter. Die Biologie Adolf Portmanns in zeitgeschichtlichem Kontext, in: Basler Zeitschrift für Geschichte und Altertumskunde, vol. 100 (2000), pp. 207–254
16. Vojta, V. Reflex rotation as a pathway to human locomotion. Z. Orthop. Ihre Grenzgeb. 1970, 108, 446–452.
18. Sanz-Esteban I, Calvo-Lobo C, Ríos-Lago M, Álvarez-Linera J, Muñoz-García D, Rodríguez-Sanz D. Mapping the human brain during a specific Vojta’s tactile input: the ipsilateral putamen’s role. Medicine (Baltimore). 2018;97(13):e0253.
19. Hok P, Opavský J, Kutín M, Tüdös Z, Kaňovský P, Hluštík P. Modulation of the sensorimotor system by sustained manual pressure stimulation. Neuroscience. 21 de 2017;348:11-22.
20. Gajewska E, Huber J, Kulczyk A, Lipiec J, Sobieska M. An attempt to explain the Vojta therapy mechanism of action using the surface polyelectromyography in healthy subjects: A pilot study. J Bodyw Mov Ther. 2018 Apr;22(2):287-292.
21. Sánchez-González, J.L.Díez-Villoria, E.; Pérez-Robledo, F.;Sanz-Esteban, I.; Llamas-Ramos, I.; Llamas-Ramos, R.; Fuente, A.d.l.;Bermejo-Gil, B.M.; Canal-Bedia, R.;Martín-Nogueras, A.M. Synergy of Muscle and Cortical Activation through Vojta Reflex LocomotionTherapy in Young Healthy Adults: A Pilot Randomized Controlled Trial. Biomedicines 2023, 11, 3203. https://doi.org/10.3390/ biomedicines11123203
22. Martínek, M.; Pánek, D.; Nováková, T.; Pavl ˚u, D. Analysis of Intracerebral Activity during Reflex Locomotion Stimulation According to Vojta’s Principle. Appl. Sci. 2022, 12, 2225.
23. Sanz-Esteban, I.; Cano-de-la-Cuerda, R.; San-Martín-Gómez, A.; Jiménez-Antona, C.; Monge-Pereira, E.; Estrada-Barranco, C.; Serrano, J.I. Cortical activity during sensorial tactile stimulation in healthy adults through Vojta therapy. A randomized pilot controlled trial. J. NeuroEng. Rehabil. 2021, 18, 13.
Nuestro cuerpo es el ultimo libro de JL Arsuaga (Destino 2023) que aborda la anatomía músculo-esquelética desde un enfoque cercano en un hábil ejercicio de comparación con otras especies y el registro fosil. Se trata de un libro para todos los públicos; un viaje por nuestra anatomía visible, a través de las obras de los grandes escultores, pintores, filosofos y científicos de todos los tiempos.
En este ameno paseo por la historia del arte se invita al lector a su exploración y a tomar conciencia de la obra de arte que es el cuerpo que habita y como funciona la maquinaria que transporta nuestros deseos de relacionarnos y explorar el mundo que nos rodea.
Una obra que también interpela aquellos que por nuestra profesión trabajamos en el día a día con la anatomía. Sorprende y cumple el objetivo que pretende el autor: hace pensar. Y eso es precisamente la intención de este Post, hacer una reflexión crítica sobre diferentes aspectos de esta magnífica obra.
Parte 1: El misterio del gran desarrollo de nuestro Glúteo Mayor (GM)
Este enigma se aborda en el capítulo titulado: La pelvis y las nalgas y expone las diferentes teorías que tratan de explicar por qué el GM se desarrolló de forma extraorinaria si apenas interviene en nuestra locomoción bípeda. Entre las cuatro teorías mencionadas, Arsuaga nombra la mía sobre la posición de cuclillas, publicada como ensayo científico en este Blog https://terapiavojtayevolucion.wordpress.com/2019/02/02/por-que-tenemos-el-culo-tan-grande/, y pendiente de publicación en una revista especializada. Por ello, me siento muy honrado y agradecido.
En esta primera entrega sobre Nuestro cuerpo, no pretendo barrer para casa, tan solo ofrecer mi humilde punto de vista de las teorías citadas.
Teoría dela carrera
La adaptación a la carrera de resistencia es la teoría de Lieberman (2006) para justificar el gran tamaño del GM al erigirse como una habilidad crítica en la caza de persecución. Arsuaga conincide con Lieberman en que el primero en nuestra filogenia en adquirir esta singular característica sería Homo Erectus. Dado que el GM apenas entra en juego en la marcha horizontal y se activa un poco más en la carrera (lo hace intensamente en el sprint) Lieberman postuló que esta actividad necesitaba un GM potente que ayudase a mantener el tronco enderezado evitando su colapso al correr con el tronco ligeramente inclinado hacia delante.
la inclinación del tronco no es la norma
Una teoria que plantea en mi opinión, serios inconvenientes: i) Lieberman asume que la inclinación del tronco sería la postura ideal por su eficiencia para la carrera de resistencia. Muy al contrario, la inclinación se asocia a mayor coste metabólico, de hecho, a mayor entrenamiento en la carrera de resistencia menor inclinación en el tronco. (Preece 2016).
Además, la fatiga en la carrera se relaciona con incrementos en la inclinación del tronco así como de la flexión de la cadera y rodilla, (Koblbauer 2014) que no responden a un agotamiento del GM (dada su baja participación) sino al de la verdadera musculatura enderezadora del raquis (erectores espinales) y la pelvis (isquiotibiales y abdominales). Este video del final del triaplon les dará una idea de lo que hablamos. El colapso de los erectores no puede ser compensado por el GM porque esa no es su función. https://www.youtube.com/watch?v=MTn1v5TGK_w&t=7s
La evidencia no apoya que el GM se desarrollase para compesar la inclinación del tronco en la carrera de resistencia, En cuanto a su implicación en la carrera durante la extenxión de la cadera es muy pequeña debido a que la fuerza de inercia hace casi todo el trabajo (Bramble 2004).
No obstante, es facil que pueda confundirle ver a grandes campeones de fondo correr con el tronco muy inclinado hacia delante (hasta 18º). Se trata de variaciones individuales adquiridas y fijadas de forma muy temprana y que han conseguido una alta eficiencia a pesar de tener una alineación del tronco no ideal. Aunque no tenemos estudios en estos atletas apuesto a que el GM participa menos que en otros atletas (no lo necesitan) ya que la inclinación del tronco reduce el rango de la extensión.
ii) los corredores de resistencia presentan un tamaño muy reducido de su GM en comparación con otros tipos de carrera de menor distancia y máxima velocidad, donde el GM se contre intensamente, lo cual podría hacer pensar que el sprint, con el GM como protagonista, fuese una presión de selección plausible (Bartlett 2014). Sin embargo, ser el más lento y menos resistente entre los velocistas de la selva o sabana pone en serios aprietos esta alternativa. A este respecto suele decirse, con cierta gracia, que no es necesario correr más que el depredador sino más que otro colega, como si el ataque fuese una especie de pistoletazo de salida en una carrera de100m lisos. Argumento tan ingenioso como ingénuo.
iii) el superdesarrollo del GM, al contrario de lo que sostiene Lieberman, surgió al menos un millon de años antes de Homo erectus, esto es con Australopitecus (poco adaptados para la carrera). En este sentido, la evidencia surge de la famosa pelvis de Lucy, plenamente bípeda, con sus reducidos isquiones (como los nuestros). A pesar de su corta estatura es altamente probable que no apoyase sus isquiones al adoptar una postura esencial en todos los primates: las cuclillas.
Las pruebas proceden de su índice intermebral similar al de un niño de 2 años y a la ausencia de callo oseo en los isquios (Arsuaga 2023), que avalan la necesidad de un GM superdesarrolado, tanto para mantenerse como para salir de las cuclillas en sentido vertical en lugar de hacerlo hacia la horizontal como hacen otros primates.
iv) Además, los estudios que aporta Liberman midieron la actividad del GM en la carrera en sujetos adultos, cuando hasta los 9 años no madura como patron. Biologicamente carece de sentido mantener una estrucutra metabolicamente tan costosa, como es el músculo, para una función que se toma una década en estar plenamente operativa. ¿Tendría sentido nacer con los caracteres sexuales desarrollados cuando la madurez reproductiva surge a partir de la 2ª década de la vida?
Teoría deLas cuclillas
La posición de cuclillas, tan innata y utlizada en la edad adulta por dos tercios de la población mundial, no es una postura para ponerse de pie, ya que es una bipedestación en sí misma (literalmente es una postura bípeda) en su forma plegada. Se emplea tanto para las actividades de la vida diaria (trabajo, ocio, relaciones sociales etc) como para el descanso ocasional de la bipedia vertical o desplegada en las sociedades orientales, y amplias zonas del continente sudamericano y africano.
Tambien mi teoría ha sido cuestionada. El argumento viene por el desuso o la falta de práctica en la edad adulta de las cuclillas en las sociedades occidentales, donde el GM debería experimentar una atrofia significativa que no se produce en relación a las sociedades orientales. Si su tamaño se relaciona con la posición de cuclillas ¿por qué se mantiene y no se atrofia completamente en los occidentales?
La respuesta es sencilla. Las cuclillas es una posicion innata, y por tanto, universal. Esta presente en todas las soceidades humanas desde el mismo instante que el niño comienza a caminar hasta que es domesticado hacia la silla en el inicio de la escolarización al estilo occidental. Por tanto, su desarrollo y tamaño viene predeterminado geneticamente porque cumple una función esencial: es la postura que sostiene el pilar de cualquier sociedad de primates: establecer y estrechar vínculos sociales.
Además, en occidente, su activadad aunque limitada, no se deroga nunca por completo ya que participa en múltiples actividades de la vida diaria como es levantarnos de nuestras sillas o de la cama, lo que supone una media sentadilla, o bien al subir escalones, cuestas, agacharnos y otras. Actividades que realizamos varias veces a lo largo del día, y que mantienen el trofismo transmitido por la configuración de un marcador epigenético después de reducir nuestros isquiones y dejarnos literalmente con el culo al aire.
Teoría sexual
La teoría sexual, de Desmond Morris (El mono desnudo 1967) tiene fuertes fundamentos para explicar la desigual distribución de la grasa en ambos sexos, pero no explica el tamaño del GM hasta la adolescencia. Al igual que sucede con la teoría de la carrera de Lieberman carece de sentido biológico mantener una estrucutra tan cara como el músculo sin otra función que rellenar una región que bien podría cubrirse por completo de tejido graso como pasa en el caso de las mamas humanas.
Teoría del lanzamiento de precisión
La teoría del lanzamiento de precisión para justificar el desarrollo del GM se aborda en el capítulo Golpear a distancia. Es imaginativa pero con un sustento científico muy limitado. El estudio que refiere Arsuaga (Marzke 1988) y que avala esta hipótesis mide mediante EMG la implicación del GM en ambas piernas durante el lanzamietno del pitcher de Beisbol.
El patron de lanzamiento se divide en 4 fases principales. De ellas, el GM se activa intensamente en la fase cuatro (despues de soltar la bola), en la pierna contraria al brazo de lanzamiento, y de ahí se infiere que los sujetos con el GM superdesarrollado serían más hábiles, y por tanto, tendrían más éxito en la reproducción (por su potencial cazador y luchador) transmitiendo sus genes de GM superdesarrollado a su descendencia.
Pero este estudio solo se centra en la actividad del GM de forma aislada. Así, cuando en el mismo lanzamiento, se estudia con la misma técnica (EMG) otros músculos de la pierna (grastronemios, vasto interno, recto femoral, glúteo mayor y biceps femoral), resulta que el GM no es el protagonista de la acción de frenado que desencadena el efecto latigo o «catapulta» en la fase 3 del lanzamiento (Campbell 2010).
La primera posición la ocupa, de forma destacada, los gastrocnemios, seguido del vasto interno, recto femoral y en tercer lugar, casi empatado con el bíceps femoral, se sitúa el GM. Además, el bíceps femoral presenta una contracción relativa mayor que el GM en relación a su potencial de contracción máxima. El GM es el que más se contrae después de la suleta de la bola (fase 4),y veremos porqué…
Tomado de Campbell 2010
Para qué se activa el GM si no es para hacer el efecto catapulta?
Fase 4 del lanzamiento, donde la pelvis rota internamente sobre el fémur
Arsuaga apunta en una dirección, que en mi opinión, no es la adecuada en la pag 455, dice: «cuando es el pie el que esta fijo la contracción del glúteo mayor produce una rotación interna de la cadera, de manera que gira hacia el otro pie. Y es ese movimiento el que resulta crucial al arrojar una piedra»
Es cierto que la pelvis describe un movimiento de rotación interna sobre la cadera del pie de apoyo, contrario a la mano de lanzamiento, en ese instante del lanzamiento (ver foto), pero desde la biomecánica no es posible atribuírselo al GM. Cuando el pie es punto fijo para el fémur, el GM sigue siendo rotador externo, es decir no aproxima la otra hemipelvis al pie de apoyo sino que la separaría. Sin embargo, efectivamente el GM esta muy activo al final del lanzamiento (fase 4) y vemos precisamente lo contrario, es decir, la rotación interna.
Es comprensible asumir que puesto que la cadera de apoyo esta en rotación interna y el GM esta fuertemente contraido, se establezca una asociación, pero muchas veces en biomecanica no es lo que parece. Entonces, sino es el GM ¿quien la hace? y ¿por qué esta tan contraido el GM en esa pierna de apoyo?
La respuesta esta en un complejo juego de fuerzas biomecánicas. La rotación interna para la cadera de apoyo en la fase del despeque de la bola, la producen dos grupos: los addutores, que tienen su punto fijo en el fémur y traccionan del pubis haciendo girar la pelvis en el plano transversal hacia la pierna de apoyo, lo cual efectivamente, implica una rotación interna para la cadera. Y el otro grupo muscular que imprime la rotación interna de la cadera de apoyo lo forman la cadena diagonal del oblicuo interno del lado apoyado con el oblicuo externo del lado contrario. Este giro del tronco sobre la pelvis de apoyo es sinergista de la rotación interna de la cadera que tiene el pie en apoyo.
Entonces, ¿qué hace el GM con su gran contracción en fase 4ª? controlar la amplitud de los rangos de movimiento de addutores y oblicuos, protejiendo la cadera de una mecánica luxante. ¿Es importante su función? por supuesto, pero no imprescindible ya que su déficit podría ser compensado por otros 6 músculos pelvitrocantereos, que por su acción rotadora externa de la cadera tambien cumplen la función de controlar un exceso de rotación interna.
patron maduro
Además, esta teoría presenta el mismo problema que la teoría de Lieberman sobre la carrera, y es que el lanzamiento, siendo una habilidad innata, alzanza la madurez de su patrón de coordinación (el medido en este estudio) en el entorno de los 9 años (Clemente 2009). De nuevo, carece de viabilidad biológica mantener un músculo tan desarrollado para cumplir su función óptima cerca de la 2ª década de la vida.
Así, el GM es un eslabón más en esta actividad (y no entre los más fuertes) por lo que esta teoría esta lejos de explicar su desarrollo por la función del lanzamineto de precisión. Los chimpancés también realizan lanzamientos, pero su potencia y precisión no es comparable con nuestro lanzamiento (Roach 2013).
La razón no esta en ningún grupo musuclar, y menos aún el GM, sino en la ausencia del patron de coordinación que incluye componentes cinemáticos esnciales como la flexión dorsal de la muñeca, ausente en el chimpancé. Pruebe a hacer un lazamiento con todas sus fuerzas bloqueando el movimiento de la muñeca hacía atrás (flexión dorsal) y comprobará la diferencia.
Reflexiones finales
De las teorías que tratan de explicar el gran desarrollo del GM solo las cuclillas supera, en mi opinión (y no porque sea mía), los mínimos exigibles para ser considerada como teoría sólida desde un enfoque científico, ya que ofrece una respuesta funcional inmediata (no en décadas) a los cambios estructurales que surgieron con la adopción de la locomoción bípeda. Sobre el surgimiento del GM Arsuaga se pregunta «si el glúteo mayor lleva a cabo en los seres humanos alguna función que no exista en los grandes simios y desde cuando la realiza». La evidencia apunta que el GM no cambio para dotarnos de una nueva función sino para mantener estable el marco postural donde se producen las relaciones sociales en los primates: las cuclillas. En otras palabras, cambió para mantener lo mismo.
La disminución de los isquiones al caer drasticamente la tracción que los isquistibiales ejercían en su inserción isquíatica debido a la nueva postura ortógrada provocó que los isquiones quedasen en el aire en la postura de sociabilización mas extendida entre los primates de antes y de ahora (humanos incluidos).
Esta situación provocó el gran desarrollo del GM, llevándole a variar la composición de sus fibras musculares hacia un predominio de las antigravitatorias tipo 1 (McAndrew 2006, O`Neill 2017). A la función extensora de la cadera, mínima en la marcha, y moderada en la carrera de fondo (para la cual no necesita este tamaño ni estrucutra), se le sumó una aún mas importante desde el punto de vista social, como es sostener la posición de cuclillas por largas horas de vida social e iniciar el despege en sentido vertical, y no en horizontal, como hacen los chimpancés y todos los simios.
En occidente esta teoría puede resultar exótica porque hemos olvidado desde los 3 años, (por imposición cultural) su enorme valor de cohesión social que hemos compensado por las prótesis que son las sillas, y también para nuestra salud postural a nivel del miembro inferior y de nuestra culumna lumbar.
Lo que confunde a muhos investigadores es que el GM participe en mayor o menor medida en diferentes actividades, y de ahí a crear una Just so story (historia ficticia e imaginativa) el paso es muy corto y tentador. Para terminar, me sumo al consejo de Arsuaga de que no se lleve la silla a la playa, pero si la silla ya es una extensión de su trasero, quizá sea temerario para su salud empezar de golpe sin un periodo de entrenamiento previo. Le propongo iniciarse con media hora sentado en el suelo como mejor prefiera e intente por unos instantes ponerse en cuclillas y hacer algo en esta posición como leer o remover la arena.
Esta incómoda y corta experiencia no va a mejorar su salud, pero tal vez le ayude a tomar conciencia de las limitaciones que ha ido adquiriendo desde su infancia cuando las cuclillas le costaban el mismo esfuerzo que respirar.
Hasta aquí, mi aportación reflexiva sobre este apasionante tema que aborda mi admirado y amigo el profesor Arsuaga…y tú qué piensas?
Bibliografia
Lieberman DE, Raichlen DA, Pontzer H, Bramble DM, Cutright-Smith E. The human gluteus maximus and its role in running. J Exp Biol. 2006;209(Pt 11):2143-55.
Bramble DM, Lieberman DE. Endurance running and the evolution of Homo. Nature. 2004 Nov 18;432(7015):345-52.
Preece SJ, Mason D, Bramah C. How do elite endurance runners alter movements of the spine and pelvis as running speed increases? Gait Posture. 2016 May;46:132-4. doi: 10.1016/j.gaitpost.2016.03.011. Epub 2016 Mar 19. PMID: 27131190.
Koblbauer IF, van Schooten KS, Verhagen EA, van Dieën JH. Kinematic changes during running-induced fatigue and relations with core endurance in novice runners. J Sci Med Sport. 2014 Jul;17(4):419-24. doi: 10.1016/j.jsams.2013.05.013. Epub 2013 Jun 19. PMID: 23790535.
Arsuaga JL. Nuetro cuerpo. Ed Destino 2023
Marzke MW, Longhill JM, Rasmussen SA. Gluteus maximus muscle function and the origin of hominid bipedality. Am J Phys Anthropol. 1988 Dec;77(4):519-28. doi: 10.1002/ajpa.1330770412. PMID: 3223519.
Bartlett JL, Sumner B, Ellis RG, Kram R. Activity and functions of the human gluteal muscles in walking, running, sprinting, and climbing. Am J Phys Anthropol. 2014 Jan;153(1):124-31.
Johnson MA, Polgar J, Weightman D, Appelton D. Data on the distribution of fibre types in thirty-six human muscles. An autopsy study. J Neurol Sci 1973; 18:111–129.
McAndrew D, Gorelick M, Brown JMM. Muscles within muscles: a mechanomyographic analysis of muscle segment contractile properties within human gluteus maximus. J Musculoskel Res 2006;10:23e35.
O’Neill MC, Umberger BR, Holowka NB, Larson SG, Reiser PJ. Chimpanzee super strength and human skeletal muscle evolution. Proc Natl Acad Sci U S A. 2017 Jul 11;114(28):7343-7348.
Las disfunciones del equilibrio suponen uno de los mayores retos en el proceso de neurorehabilitación de las lesiones neurológicas. La velocidad y precisión necesaria para mantener constantemente el centro de masas dentro de la base de sustentación convierten a estas reacciones automáticas en una víctima de primer orden ante un evento cerebral.
Entre los procedimientos más empleados para mejorar esta función básica en la locomoción existen una gran variedad de estímulos y técnicas entre las que destacan la realidad virtual (Rooij 2021), entrenamiento orientado a tareas (Ahmed 2021), balance reactivo (Handelzalts 2019) y tai-chi (Zheng 2021) entre las más destacadas.
2. El balance reactivo
Este post esta inspirado en la interesante entrevista realizada por Javier Sánchez Aguilar en su canal de Hemispherics con Gonzalo Varas, Kinesiologo especializado en neurorehabilitación, la cual estoy encantado de recomendar: https://www.youtube.com/watch?v=eATrGXRAVbs&t=836s
Bebé levantándose tras una caída
Si tuviera que resumir mucho el concepto de balance reactivo, sobre la que gira la charla, diría que consiste en aplicar perturbaciones en el equilibrio del paciente, (mediante dispositivos diseñados al efecto) que provocan estrategias automáticas de recuperación del equilibrio perdido, utilizando un arnés de seguridad para evitar riesgos.
Frente al cuidado que solemos tener los fisios para evitar la caída durante nuestros tratamientos este procedimiento parece revolucionario, pero si echamos un vistazo al desempeño motor del niño que ha alcanzado la marcha recientemente, las caídas son una constante en las primeras semanas o meses, pudiendo llegar este periodo de consolidación del equilibrio hasta los 3 años (Vojta 2011).
3. Las caídas en el proceso de adquisición de la marcha.
Un vistazo al neurodesarrollo del equilibrio en la marcha del bebé humano y su comparación con la marcha erguida del bebé chimpancé nos hace plantear la siguiente cuestión: ¿Por qué son tan frecuentes las caídas en el bebé humano frente al bebé chimpancé?
En el bebé humano la práctica febril de la marcha se considera la responsable de los ajustes posturales que progresivamente van mejorando la coordinación del patron. Caerse, por tanto, sería parte necesaria del proceso de aprendizaje, por lo que el balance reactivo que explica Gonzalo Varas tiene, en este contexto, todo el sentido. Asi, la cuestión podría darse por zanjada: practicar el desequilibrio mejora el equilibrio ¿acaso cabe otra explicación a la progresiva mejora en calidad y seguridad en la marcha de la cría humana?
3.1 Equilibrio comparado. Si consideramos que las caídas en el bebé chimpancé que práctica la marcha erguida son muy infrecuentes ¿a qué se debe esta diferencia? la hipótesis de la neotenia o infancia extendida (Gould 2010) apunta al tiempo empleado para alcanzar la madurez en sus respectivas locomociones: de 4 a 5 meses para la cuadrúpeda del chimpancé, por los 3 años para la bípeda humana. Mientras que la bipedia transitoria que práctica el bebé chimpancé lo hace desde la madurez de su patrón locomocor cuadrúpedo, el humano lo hace desde la inmadurez del propio.
¿Qué aspectos caracterizan la inmadurez de la marcha en el bebé humano? base de sustentación ancha, brazos en asa, pelvis antevertida (delatada por el deficit de activación abdominal), movimientos laterales en el tronco (extensión axial deficitaria), contacto plano del pie, inestabilidad, parámetros espacio-temporales alterados, entre otros (Ivanenko 2007).
En cambio, en la marcha erguida del bebé chimpancé las diferencias cinesiológicas respecto a su marcha erguida de adultos se reducen a un incremento leve de la extensión de las articulaciones de cadera y rodillas, lo que redunda en mayor ahorro energético (Kimura 1996). ¿Qué importancia tiene ésto para el caso que nos ocupa? que la madurez del patron locomotor y el equilibrio van de la mano y son interdependientes.
4. Los secretos del equilibriohumano.
Después de un periodo de inestabilidad en la locomoción bípeda del bebe humano ¿Por qué a nosotros nos resulta tan fácil y económico mantener el equilibrio en nuestro caminar bípedo en comparación con los chimpancés?
4.1 Factores estructurales:
eje longitudinal
4.1.1. Columna vertebral erguida con el agujero magno en la base del craneo que permite que el eje longitudinal este alineado con el eje axial frente a la cifótica del chimpancé con su agujero magno situado en la zona posterior. De esta forma, con muy poca actividad musuclar el sistema vestibular consigue una gran eficiencia de las reacciones de equilibrio.
4.1.2 Pelvis con iliacos orientados lateralmente que posibilita una actividad enderezadora del glúteo medio en el plano frontal (abductor), en lugar de la orientación dorsal del chimpancé donde el gluteo mediano es otro extensor más (Arsuaga 1998).
4.1.3 Angulo bicondiliar de nuestras rodillas que hacen que convergan hacia la línea media en valgo. Esto asegura que ambos pies se sitúen justo por debajo de la pelvis, a diferencia del chimpancé que en bipedestacion tienen los pies más separados entre sí. De esta manera, en el humano se reduce el área de sustentación, una dificultad (exige reacciones más rápidas y precisas) que se ve compensada por la alineación del eje vertical del cuerpo. Por el contrario, caminar con los pies separados como los chimpancés y los bebés humanos (en sus primeros pasos) implica mayor movilidad del centro de masas y por tanto, reacciones más amplias, lentas y costosas (Nakatsukasa 2006).
4.2 Factores funcionales.
4.2.1 Musculatura epiaxial con predominio de fibras tipo I, más resistentes a la fatiga, frente a las de tipo II del chimpancé (Oneill 2017). Esto nos permite sostener la CV en el eje axial generando de esta forma un punto de apoyo estable en la región torácica donde los chimpancés no pueden, visible por una marcada inclinación lateral durante el apoyo monopodal.
Washburn (1968) asociaba esta inclinación con la orientación dorsal de los iliacos del chimpancé. Sin embargo, Hirasaki (2004) ha demostrado que el entrenamiento temparano en la bipedia reduce al minimo estas inclinaciones laterales. La composición del tipo de fibras musculares de los grupos epiaxiales esta determinada geneticamente en cada especie en función de su tipo de locomoción (Shapiro 1988). Una composición que puede ser modificada por el entrenamiento intenso y temprano (Pette 2000, Staron 1997, Wilson 2012). De hecho, la estimulación electrica aislada de esta musculatura en pacientes post-ictus ha demostrado ser efectiva en la mejora del equilibrio (Bilek 2020).
Otro factor en contra de la hipótesis estructural la encontramos en la marcha inmadura del bebé donde puede observarse movimientos de inclinación similares en el tronco, lo cual responde a un periodo donde aún la musculatura epiaxial no puede sujetar la CV en su eje longitudinal tras liberar las manos del apoyo para caminar.
bebe en sus primeros pasos
4.2.2 Retroversión. Esta es una característica espefícica humana deribada de nuestra capacidad para mantener la CV en el eje axial en bidestación, Con ello, surgen nuevas fuerzas vectoriales en la región abdominal hacia este punto de anclaje en la CV torácica a través de las cadenas oblicuas abdominales. Los chimpancés no tienen este vector, y para mantenerse erguidos deben compensar este déficit por medio de las inclinaciones laterales y anterior del tronco, junto con la flexión de las caderas y rodillas (Thompson 2015),
4.2.3. Control postural automático. Común a todas las especies que tienen locomoción (Grillner 1981). El nivel de procesamiento automático esta controlado por estructuras subcorticales (Región locomotora mesencefalica, Ganglios basales y cerebelo) y medulares, incluyendo los generadores de patrones centrales (GPC).
5. El equilibrio desde la Locomoción Refleja de Vojta.
Diferentes estudios apuntan la efectividad de la Locomoción Refleja en la mejora del equilibrio en el paciente neurológico (Min 2017, Epple 2020, Perales 2021, Carratala 2022) a pesar de que ninguno de sus patrones práctica el desequilibrio. ¿Como se explica? Por que actúa directamente sobre los tres factores funcionales implicados en el equilibrio de la marcha bípeda madura.
Patrón innato de la reptación refleja
Así, la extensión axial de la CV, y la retroversión de la pelvis estan presentes en los tres patrones de la LR, con un control postural automático espefícico en cada patrón (Vojta 1991). Estos patrones se activan en posiciones en decúbito, muy estables, donde las perturbaciones del equilibrio son mínimas. Sin embargo, paradojicamente, su activación tiene una potente influencia sobre las reacciones de equilibrio en bipedestación y marcha.¿Como es posible?
Porque se trata de programas motores innatos de coordinación cuadrúpeda con su específico control postural automático, cuyos núcleos de coordinación neural son comunes en ambos tipos de locomoción, la cuadrúpeda y la bípeda (Zehr 2005, Diezt 2011). Así se explica la transferencia del control automático a la marcha bípeda. Si… muy anti-intuitivo, muy loco.
6. Consideraciones finales.
La maduración neuro-cinesiológica y el equilibrio van de la mano en el neurodesarrollo. La pulsión de explorar el entorno lleva implícito el ensayo-error de las propias habilidades innatas hasta alcanzar su desarrollo óptimo. Es entones, cuando el equilibrio de nuestra singular forma de locomoción alcanza su máxima cota de eficiencia. En la patología neurológica del paciente adulto hay evidencia de que las reacciones automáticas de equilibrio deben ser abordadas igualmente de la forma mas automática posible como hace el balance reactivo. La locomoción refleja, ademas de su naturaleza intrínsecamente automática aborda los factores funcionales implicados en la maduracion del equilibrio que siempre son deficitarios en el paciente neurológico. Por esto, una combinación de estas intervenciones podría ser una interesante linea de investigación a considerar.
Bibliografia
Ahmed U, Karimi H, Amir S, Ahmed A. Efectos del entrenamiento intensivo del tronco multiplanar junto con ejercicios de doble tarea sobre el equilibrio, la movilidad y el riesgo de caídas en pacientes con accidente cerebrovascular: un ensayo controlado aleatorio. J Int Med Res. 2021 noviembre; 49 (11): 3000605211059413. doi: 10.1177/03000605211059413. PMID: 34812070; PMCID: PMC8647262.
Arsuaga JL, Martinez Mendizabal I. La especie elegida. Temas de Hoy. 1998
Bilek F, Deniz G, Ercan Z, Cetisli Korkmaz N, Alkan G. The effect of additional neuromuscular electrical stimulation applied to erector spinae muscles on functional capacity, balance and mobility in post-stroke patients. NeuroRehabilitation. 2020;47(2):181-189. doi: 10.3233/NRE-203114. PMID: 32741788.
Carratalá-Tejada M, Cuesta-Gómez A, Ortiz-Gutiérrez R, Molina-Rueda F, Luna-Oliva L, Miangolarra-Page JC. Reflex Locomotion Therapy for Balance, Gait, and Fatigue Rehabilitation in Subjects with Multiple Sclerosis. J Clin Med. 2022 Jan 23;11(3):567. doi: 10.3390/jcm11030567. PMID: 35160020; PMCID: PMC8836377.
de Rooij IJM, van de Port IGL, Punt M, Abbink-van Moorsel PJM, Kortsmit M, van Eijk RPA, Visser-Meily JMA, Meijer JG. Effect of Virtual Reality Gait Training on Participation in Survivors of Subacute Stroke: A Randomized Controlled Trial. Phys Ther. 2021 May 4;101(5):pzab051. doi: 10.1093/ptj/pzab051. PMID: 33594443; PMCID: PMC8122468.
Dietz V. Quadrupedal coordination of bipedal gait: implications for movement disorders. J Neurol. 2011 Aug;258(8):1406-12.
Epple, C., Maurer-Burkhard, B., Lichti, M. et al. Vojta therapy improves postural control in very early stroke rehabilitation: a randomised controlled pilot trial. Neurol. Res. Pract.2, 23 (2020). https://doi.org/10.1186/s42466-020-00070-4
Grillner S (1981) Control of locomotion in bipeds, tetrapods and fish. In: Brookhart JMaMVB (ed) Handbook of physiology. The nervous system. Motor control. American Physilogy Society, New York, pp 1179–1236.
Gould Jay Stephen. Ontogenia y filogenia. La ley fundamental biogenetica.Editorial critica 2010.
Handelzalts S, Kenner-Furman M, Gray G, Soroker N, Shani G, Melzer I. Effects of Perturbation-Based Balance Training in Subacute Persons With Stroke: A Randomized Controlled Trial. Neurorehabil Neural Repair. 2019 Mar;33(3):213-224. doi: 10.1177/1545968319829453. Epub 2019 Feb 15. PMID: 30767613.
Hirasaki E, Ogihara N, Hamada Y, Kumakura H, Nakatsukasa M. Do highly trained monkeys walk like humans? A kinematic study of bipedal locomotion in bipedally trained Japanese macaques. J Hum Evol. 2004 Jun;46(6):739-50. doi: 10.1016/j.jhevol.2004.04.004. PMID: 15183673.
Ivanenko YP, Dominici N, Lacquaniti F. Development of independent walking in toddlers. Exerc Sport Sci Rev. 2007 Apr;35(2):67-73. doi: 10.1249/JES.0b013e31803eafa8. PMID: 17417053.
Kimura T. Centre of gravity of the body during the ontogeny of chimpanzee bipedal walking. Folia Primatol (Basel). 1996;66(1-4):126-36. doi: 10.1159/000157190. PMID: 8953755.
Min Soo Sonb, Do Hee Junga, Joshua (Sung) H. Youa,∗, Chung Hwi Yic, Hye Seon Jeonc. Effects of dynamic neuromuscular stabilization on diaphragm movement, postural control, balance and gait performance in cerebral palsy. NeuroRehabilitation 41 (2017) 739–746.
Nakatsukasa M, Hirasaki E, Ogihara N. Energy expenditure of bipedal walking is higher than that of quadrupedal walking in Japanese macaques. Am J Phys Anthropol. 2006 Sep;131(1):33-7. doi: 10.1002/ajpa.20403. PMID: 16485295.
O’Neill MC, Umberger BR, Holowka NB, Larson SG, Reiser PJ. Chimpanzee super strength and human skeletal muscle evolution. Proc Natl Acad Sci U S A. 2017;114(28):7343-7348. doi:10.1073/pnas.1619071114
Perales Lopez L, Palmero NV, Ruano LG, San Leon Pascual C, Orile PW, Down AV, Gor Garcia-Fogeda MD, Toré S. The implementation of a reflex locomotion program according to Vojta produces short-term automatic postural control changes in patients with multiple sclerosis. J Bodyw Mov Ther. 2021 Apr;26:401-405. doi: 10.1016/j.jbmt.2021.01.001. Epub 2021 Jan 9. PMID: 33992274.
Pette D, Staron RS. Myosin isoforms, muscle fiber types, and transitions. Microsc Res Tech. 2000;50(6):500-509. doi:10.1002/1097-0029(20000915)50:6<500::AID-JEMT7>3.0.CO;2-7
Shapiro LJ, Jungers WL. Back muscle function during bipedal walking in chimpanzee and gibbon: implications for the evolution of human locomotion. Am J Phys Anthropol. 1988;77(2):201-212. doi:10.1002/ajpa.1330770208.
Staron RS. Human skeletal muscle fiber types: delineation, development, and distribution. Can J Appl Physiol. 1997;22(4):307-327. doi:10.1139/h97-020
Thompson NE, Demes B, O’Neill MC, Holowka NB, Larson SG. Surprising trunk rotational capabilities in chimpanzees and implications for bipedal walking proficiency in early hominins. Nat Commun. 2015 Oct 6;6:8416.
Vojta V, Schweizer E. El descubrimiento de la motricidad ideal. Ediciones Morata 2011.
Washburn SL. 1968. The study of human evolution. Eugene, OR: Oregon State System of Higher Education.
Wilson JM, Loenneke JP, Jo E, Wilson GJ, Zourdos MC, Kim JS. The effects of endurance, strength, and power training on muscle fiber type shifting. J Strength Cond Res. 2012;26(6):1724-1729. doi:10.1519/JSC.0b013e318234eb6f
Zehr, E.P. Neural control of rhythmic human movement: the common core hypothesis. Exerc. Sport Sci. Rev. 33:54Y60, 2005.
Zheng X, Wu X, Liu Z, Wang J, Wang K, Yin J, Wang X. The Influences of Tai Chi on Balance Function and Exercise Capacity among Stroke Patients: A Meta-Analysis. Evid Based Complement Alternat Med. 2021 Feb 24;2021:6636847. doi: 10.1155/2021/6636847. PMID: 33708256; PMCID: PMC7932789.
Plantear como nuevo paradigma un enfoque neurorehabilitador publicado por primera vez en los setenta puede resultar paradójico (Vojta 1972). Hablo de las Locomociones reflejas (LR) de Vojta o activación de los patrones genéticos de la locomoción.
En este momento es posible que ya no quiera seguir leyendo este post, a causa de los prejuicios acerca de lo que cree conocer sobre esta terapia…
Pero si decide continuar, estoy seguro de que se le abrirán nuevas perspectivas que pueden enriquecer su visión del tema, y tal vez pueda considerar la existencia de un nuevo concepto de abordaje en la neurorehabilitación del paciente adulto desde la genética de la motricidad humana.
2. Un cambio conceptual
Este Post no plantea un cambio del paradigma neurorehabilitador (a nivel motor) basado principalmente en el entrenamiento intensivo de la función con diferentes técnicas, y dispositivos tecnológicos (robots, realidad virtual, Terapia restrictiva, doble tarea, etc). El entrenamiento orientado a tareas es eficaz (Chan 2006) para mejorar el desempeño funcional en actividades diseñadas por terapeutas ocupacionales.
Entrenamiento asistido de la función de locomoción
Lo que a continuación se plantea es un cambio conceptual, que implica una invitación a la reflexión para poner en un contexto global el paradigma funcional.
Los cambios conceptuales nunca son fáciles, y aunque parezca difícil de creer no dependen exclusivamente de la evidencia científica, sino del marco mental dominante. Para que un nuevo concepto sea aceptado influye, y mucho, que el «nuevo» esté, si no en la misma línea de pensamiento, en una admisible y coherente con los conocimientos previos; o por lo menos, que no vaya en una dirección inasumible por las ideas ya instauradas.
En relación al paradigma actual, la LR es precisamente eso, inasumible, ya que no encaja con los conocimientos previos basados en el entrenamiento de la función. Las LR de Vojta no trabajan actividades funcionales y no son auto-iniciadas, dos premisas que, por sí mismas descartan este enfoque según el paradigma imperante.
3.Conflicto cognitivo
Pero la resistencia a aceptar la existencia de una vía genética de la motricidad, también en humanos, se mantiene mas allá de lo que la fisiología del desarrollo motor haya constatado (Grilner 1985, Ritter 2000, Diezt 2001). Los prejuicios son tan prevalentes que ni siquiera la generación de un fuerte conflicto cognitivo como es la demostración de los efectos de las LR sobre el propio cuerpo del más exigente y experto de los clínicos puede superar esta barrera.
Adulto activada en patrón de Reptación refleja
¿Imagina que después de estar posicionado en una postura concreta, donde le presionan distintas zonas de su cuerpo durante unos minutos, se pone de pie y su cuerpo comienza a caminar de forma automática sin que usted haya iniciado este movimiento?
Una experiencia de este tipo debería cuanto menos provocar una serie de preguntas que hiciesen tambalear las ideas previas sobre lo que cree conocer en relación a la marcha y su fisiología.
Sin embargo, lo que me encuentro habitualmente es a colegas que tratan de buscar justificaciones imaginativas para auto-convencerse que lo que acaba de suceder tiene una explicación; sugestión, hipnosis, inducción guiada y otras cosas similares.
Es el llamado sesgo de confirmación. De hecho, si las nuevas ideas no encajan con nuestra línea de pensamiento se activan áreas cerebrales asociadas a la detección de errores y la inhibición de respuestas (Fugellsang 2005). Entonces la posición natural es defenderse ante esta «agresión», bien ignorando, negando, o combatiendo al nuevo concepto.
Además, asumir un nuevo concepto de forma consecuente exige que éste se entienda bajo el prisma del conocimiento previo, ya que nuestra mente necesita rellenar y allanar el vacío que separa el conocimiento previo y el nuevo. De alguna manera es necesario establecer un vinculo que ayude en la transición del marco mental y reduzca las fricciones del conflicto cognitivo que va a producirse.
Suponiendo que se consigue superar el rechazo inicial y encontramos cierta receptividad a escuchar y experimentar las LR, todavía hay que enfrentar tres puntos relevantes según el modelo de Posner (1982) sobre la asimilación de un nuevo concepto: que sea (1)inteligible, (2)coherente y (3)predictivo.
3.1 Inteligible
¿Cómo se puede hacer inteligible una transición entre el paradigma basado en el entrenamiento de la función a otro basado en la activación de la genética de la motricidad humana?
(a) Partiendo desde un enfoque más amplio que abarque la biología evolutiva, la ontogenia y la filogenia de nuestra locomoción bípeda y la cinesiología esencial del miembro superior que facilite comprender el contexto evolutivo en el que se desarrollo nuestra motricidad especifica, así como la relación de éste proceso con el posible origen de las zonas de activación de las LR. Solo así podrá entenderse la naturaleza genética de nuestra motricidad como especie. Para ampliar información ver: teoria-neuro-plastica-de-la-evolucion-del-bipedalismo-humano/
Bajo este enfoque la función se entiende como un subproducto derivado de la interacción de los programas motores genéticos con el ambiente. Los humanos como cualquier otra especie, no entrenamos función alguna, si no que esta es consecuencia y no causa de la interacción.
Por tanto, la función es enormemente adaptativa y flexible según el entorno que se habita. Lo que hoy día entendemos en occidente como AVD nada tienen que ver con las AVD de una población amazónica, Inuit, o oriunda de la sabana africana. Sin embargo, todas estas poblaciones comparten la misma genética en lo relativo a su comportamiento motor y es su interacción con su hábitat el que determinará qué tipo de actividades son funcionales y cuales no.
La función es dependiente de la interacción de los programas motores genéticos con el entorno
Por supuesto, la continua interacción con el medio mejorará las competencias en cada función desempeñada, como sucede en cualquier persona que se enfrenta a un nuevo reto motor como es conducir, montar en bici o subir a una palmera. Así, la función se puede ver como un medio, no un fin. Un medio que se sirve de los patrones motóricos genéticos para alcanzar su objetivo.
(b)Otro aspecto esencial para hacer inteligible este concepto pasa por la difusión de las monografias (Vojta 1972, Vojta 1992, Orth 2019) que muestran como acceder a estos patrones universales (genéticos) de forma sistemática y a cualquier edad por medio de la aplicación de las LR. La fácil reproducibilidad de los patrones de LR permite a otros clínicos siguiendo las pautas marcadas en estos trabajos replicar resultados similares en sujetos sanos (Pavlu 2000, Perales 2013, Gajewska 2018). Por sus características el Principio Vojta permite diseñar estudios ECA a doble ciego.
3.2 Coherente
Sobre la base del punto anterior, el conflicto cognitivo debería ir reduciéndose sustentado en dos aspectos esenciales: (1) el empírico, apoyado en la experimentación de las sensaciones y efectos de las LR sobre uno mismo y en terceros; y (2) el científico, sustentada en los estudios sobre las estructuras neurales implicadas, sus efectos neuromoduladores (Hok 2017, Hok 2019, Sanz 2018) y clínicos en pacientes (Backstrom 2000, Kanda 2004, Epple 2020, Perales 2021).
Este punto del proceso de asimilación del nuevo concepto debería ir acompañado de preguntas donde se busca contrastar ambos modelos buscando incoherencias y compatibilidades con el paradigma actual y otros conocidos.
Pregunta 1: El entrenamiento funcional funciona?
La evidencia es clara: funciona. El entrenamiento intensivo de una una función como la locomoción mejora el rendimiento (Munari 2018, Globas 2012). Lo mismo pasa con el Miembro superior (Kwakkel 2015, Bertani 2017). El axioma es simple, lo que sirve para mejorar el rendimiento físico y la adquisición de nuevas habilidades en la población sana, también lo es (con las adaptaciones oportunas) para la población con afectación motorica de origen neurológico.
La explicación del éxito parece clara: la repetición constante. Como estrategia, la repetición parece estar más cerca del aprendizaje que del innatismo. En consecuencia, se podría presuponer un carácter no genético de patrones básicos como la marcha. Sin embargo, la repetición no constituye un factor excluyente del innatismo, sino que forma parte integral de él (Vojta 2011). La pulsión de la repetición o ensayo-error constituye, con la imitación. las estrategias innatas para el afinamiento (coordinación) y acondicionamiento de todas las habilidades motrices y cognitivas, sean de carácter innato o fruto del aprendizaje (Mora 2014).
Bebé «entrenando» la marcha
Además de añadir la variable intensidad ¿tiene margen de mejora el entrenamiento funcional como estrategia de neurorehailitación? La combinación con otras técnicas como la neuromodulación, imaginería, realidad virtual y otras así lo sugieren.
Sin embargo, aún no se ha explorado suficientemente una combinación de enorme potencial como es disponer de un acceso directo a los programas motóricos genéticos específicos de nuestra especie. Los estudios realizados con adultos apuntan que una activación de esta naturaleza permite a los pacientes disponer de éstos patrones para utilizarlos inmediatamente de forma automática, y por tanto, sin esfuerzo (Perales 2021, Epple 2020).
Al uso automático de la función que acompaña la post activación de las LR también se le podría llamar «entrenamiento» no consciente; del mismo modo podría decirse del bebé, que una vez tiene disponible el patrón genético de la marcha se entrega febrilmente a «entrenarla» por interminables horas.
Bajo este prisma el «entrenamiento» de la función pasa a un segundo plano, puesto que surge de forma espontánea (sin pretenderlo conscientemente) en la interacción de la persona con su realidad física y social.
Pregunta 2: ¿la marcha es un producto de la genética? ¿del aprendizaje? ¿de ambos?
No debemos olvidar que nuestra locomoción bípeda no es más que la expresión de una modificación en el programa genético cuadrúpedo/arborícora que emergió en un contexto ambiental determinado hace más de 5 Millones de años (Arsuaga 1998).
A pesar de que el contexto ambiental donde surgió ha cambiado, el programa bípedo continuó siendo seleccionado por su enorme valor adaptativo. Entonces el programa bípedo fue asimilado en nuestro genoma, junto con las modificaciones estructurales para este tipo de locomoción.
En cuanto a la pregunta, la respuesta combinada (genética + ambiente) parece la más lógica, aunque Shumway-Cook (1995) considera la marcha como un producto de la experiencia. Tal vez el concepto de supresión de experiencia de movilidad en el primer año de vida, puesta en practica por distintas poblaciones tribales nos brinde una respuesta. Como describe Eibesfeldt (1984):
«Se considera que un patrón es propio de la especie cuando aunque se evite su experiencia de un modo establecido, ese patrón especifico aparece en el momento adecuado de su maduración.»
En otras palabras, impedir la marcha en un niño/a sano (por cualquier medio físico) durante su neurodesarrollo no evita que esta aparezca a su edad una vez que es liberado de las restricciones.
La demostración empírica de que hablamos de un patrón 100% genético procede de la aplicación de las LR. Que las LR desencadenan el patrón de marcha sin auto iniciación voluntaria puede comprobarse en cualquier sujeto adulto sano. https://youtu.be/jL2J3ezNVWg
Secuencia real de activación del patrón de la 1ª posición de la LR en persona sana. El patrón finaliza con la marcha refleja
Pregunta 3: ¿Qué aporta la activación genética que no puede ofrecer al entrenamiento funcional?
Para responder a la pregunta permítame emplear una metáfora informática. Si su ordenador presenta un fallo del sistema operativo, éste dispone de algoritmos para tratar de superarlo y continuar operando con más o menos dificultades. Pero seguro que usted conoce otra posibilidad que habrá utilizado en más de una ocasión: reiniciar el sistema para volver a cargar los parámetros originales, los que vienen predeterminados de fabrica.
Llevando esta metáfora a la cuestión planteada, la activación de las LR representaría el «reinicio» del sistema locomotor para cargar los elementos cinesiológicos que se han perdido o que están alterados por la lesión junto con el control postural automático inherente a toda función. Obviamente la magnitud del daño condicionará las posibilidades de evocación de los patrones parciales disfuncionales.
Pregunta 4: El paradigma genético de activación refleja puede sustituir al funcional en el proceso de neruorehabilitación?
Platear una relación dicotómica entre ambos paradigmas no tiene sentido, puesto que genética y función están interconectados, ya que tanto locomoción como función manual son la resultante del programa motor genético.
En el caso de la marcha la intervención a nivel genético mediante la activación de los patrones de las LR, solo pueden expresarse a través de la función de locomoción, por tanto, la práctica posterior de la marcha bípeda es condición sine qua non para valorar el grado de asimilación de los componentes cinesiológicos ideales activados de forma refleja (Laufens 1999). Este es el motivo por el cual no se plantea un cambio de paradigma, (no hay contradicción) si no algo de mucho más calado y complejo, como es un cambio conceptual.
En la actualidad, el entrenamiento intensivo de la marcha aborda este problema ignorando que existe una forma de acceso a los programas motores ideales.
3. 3 Predictivo
Items de la OP que se relacionan con las LR.
Este nuevo concepto debería contribuir a explicar de un modo clarificador y diferente las disfunciones de la marcha y del MS en el adulto neurológico, tomando como referencia de la organización y coordinación de nuestra motricidad la ontogénesis postural (OP).
Este es el periodo critico para el ensamblaje de todos los patrones específicos de nuestra especie a lo largo del primer año de vida. No debemos olvidar que estos patrones serán los que usaremos el resto de nuestra vida (Vojta 2011).
Bajo el prisma de la OP como base del razonamiento clínico en adultos (Kobesova 2020), el fisioterapeuta podrá resolver (junto con el conocimiento de la patología) el diagnóstico funcional de cualquier alteración desde la cinesiología del neurodesarrollo aplicada al paciente adulto. Los test validados a nivel internacional serán necesarios para registrar de forma objetiva la validez de este nuevo concepto.
En la actualidad hablar de la existencia de una genética de la motricidad humana suena para muchos colegas a ciencia ficción, y aún lo es más relacionar la «vieja» terapia Vojta con un instrumento de acceso a dicha motricidad. A pesar de la creciente evidencia, este hecho continua siendo visto de forma marginal, y mucho me temo que el marco metal dominante no aceptará fácilmente la inclusión de las LR sin antes hacer la debida digestión que supone comprender en toda su dimensión sus mecanismos neurofisiológicos de acción y no solo sus resultados clínicos por positivos que puedan ser.
No será fácil, pero si inevitable que la ciencia continúe avanzando hacia el modelo neurorehabilitador más eficiente, que irá desechando unos procedimientos e integrando otros nuevos diseñados a la medida de cada paciente. Entre estas técnicas, sin duda, el papel de la genética será cada vez más relevante (como esta sucediendo en medicina) y no solo en el ámbito de la neurogénesis, sino también en la activación de los programas motóricos originales de nuestra especie que ayuden a la más eficiente reconexión entre las neuronas que controlan nuestra motricidad.
En este aspecto las LR tendrán mucho que aportar, aunque este proceso no será tan rápido como desearíamos. Queda mucho trabajo e investigación por hacer, pero a fin de cuentas así se ha escrito la historia de nuestra joven especialidad dentro de la fisioterapia.
Bibliografia
Arsuaga JL, Martinez Mendizabal I. La especie elegida. Temas de Hoy.1998.
Backstrom B. Vojta self-training: experiences of six neurologically impaired people. Physiotherapy. 2000;86(11):567-74.
Bertani R, Melegari C, De Cola MC, Bramanti A, Bramanti P, Calabrò RS. Effects of robot-assisted upper limb rehabilitation in stroke patients: a systematic review with meta-analysis. Neurol Sci. 2017 Sep;38(9):1561-1569. doi: 10.1007/s10072-017-2995-5. Epub 2017 May 24. PMID: 28540536.
Chan DY, Chan CC, Au DK. Motor relearning programme for stroke patients: a randomized controlled trial. Clin Rehabil. 2006 Mar;20(3):191-200. doi: 10.1191/0269215506cr930oa. PMID: 16634338
Dietz V, Fouad K, Bastiaanse CM. Neuronal coordination of arm and leg movements during human locomotion. Eur J Neurosci. 2001 Dec;14(11):1906-14.
Eibl-Eibesfeldt I. (1984). Die Biologie des menschlichen Verhaltens, Piper München Zürich.
Epple C., Maurer-Burkhard B., Lichti M. et al. Vojta therapy improves postural control in very early stroke rehabilitation: a randomised controlled pilot trial. Neurol. Res. Pract. 2,23 (2020). https://doi.org/10.1186/s42466-020-00070-4.
Francisco Mora. 2014. ¿Cómo funciona el cerebro? Alianza editorial.
Gajewska E, Huber J, Kulczyk A, Lipiec J, Sobieska M. An attempt to explain the Vojta therapy mechanism of action using the surface polyelectromyography in healthy subjects: A pilot study. J Bodyw Mov Ther. 2018 Apr;22(2):287-292.
Grillner S (1981) Control of locomotion in bipeds, tetrapods and fish. In: Brookhart JMaMVB (ed) Handbook of physiology. The nervous system. Motor control. American Physilogy Society, New York, pp 1179–1236.
Grillner S, Wallén P: Central pattern generators for locomotion, with special reference to vertebrates. Annu Rev Neurosci, 1985, 8: 233–261.
Hok P, Opavský J, Kutín M, Tüdös Z, Kaňovský P, Hluštík P. Modulation of the sensorimotor system by sustained manual pressure stimulation. Neuroscience. 21 de 2017;348:11-22.
Hok P, Opavský J, Labounek R, et al. Efectos diferenciales de la estimulación por presión manual sostenida según el sitio de acción. Frente Neurosci;2019; 13: 722. (2019) doi: 10.3389 / fnins.2019.00722
Husárová, R. The vojta approach in adult patients. Rehabilitacia. 2005;42(3).
Kanda T, Pidcock FS, Hayakawa K, Yamori Y, Shikata Y. Motor outcome differences between two groups of children with spastic diplegia who received different intensities of early onset physiotherapy followed for 5 years. Brain Dev 2004;26(2):118-26.
Kobesova A, Davidek P, Morris C, Andel R, Maxwell M, Oplatkova L, Safarova M, Kumagai K, Kolar P. Functional postural-stabilization tests according to Dynamic Neuromuscular Stabilization approach: Proposal of novel examination protocol, Journal of Bodywork and Movement Therapies, 24(3) 2020: 84-95, ISSN 1360-8592, https://doi.org/10.1016/j.jbmt.2020.01.009.
Kwakkel G, Veerbeek JM, van Wegen EE, Wolf SL. Constraint-induced movement therapy after stroke. Lancet Neurol. 2015 Feb;14(2):224-34. doi: 10.1016/S1474-4422(14)70160-7. PMID: 25772900; PMCID: PMC4361809.
Laufens G, Poltz W, Buchstein G, Schmiegelt F, Stempski S. [Improvement in locomotion by combined treadmill/Vojta physiotherapy applied to selected MS patients] [Article in German]. Phys Rehab Kur Med. 1999; 9: 187-189.
Laufens G, Seitz S, Staenicke G, Fachbereich. Fundamentos biológicos comparativos respecto a la locomoción innata en particular respecto a la “reptación refleja” según Vojta. Fisioterapia;43. Jg (5/1991): 448-156.9.
Munari D, Pedrinolla A, Smania N, Picelli A, Gandolfi M, Saltuari L, Schena F. High-intensity treadmill training improves gait ability, VO2peak and cost of walking in stroke survivors: preliminary results of a pilot randomized controlled trial. Eur J Phys Rehabil Med. 2018 Jun;54(3):408-418. doi: 10.23736/S1973-9087.16.04224-6. Epub 2016 Aug 30. PMID: 27575015.
Pavlu D, Véle F, HavlíˇckováL. Electromyography and kinesiological analysis of Vojta´s therapeutical principle. Rehab Fyzil. 2000;7:34-9.
Perales L, Pérez AM, Atim MA, Varela E. Efecto de la terapia Vojta en la rehabilitación de la marcha en dos pacientes adultos con daño cerebral adquirido en fase tardía. Fisioterapia. 2009;31:151—62.
Perales Lopez L, Valdez Palmero N, Garcia Ruano L, San Leon Pascual C, White Orile P, Vegue Down A, Gor Garcia-Fogeda MD, Toré S. The implementation of a reflex locomotion program according to Vojta produces short-term automatic postural control changes in patients with multiple sclerosis. J Bodyw Mov Ther. 2021. 26: 401-405. ISSN 1360-8592, https://doi.org/10.1016/j.jbmt.2021.01.001.
Perales-López L, Fernández-Acenero M.J. ¿Es transferible la terapia de locomoción refleja a una plataforma de teleneurorrehabilitación en el tratamiento del paciente adulto? Rehabilitación. 2013;47(4):205-212.
Posner G.J, et al (1982). Accommodation of a scientific conception. Toward a theory of conceptual change. Science education, 66(2), 211-227.
Ritter M. Die Biologie Adolf Portmanns in zeitgeschichtlichem Kontext, in: Basler Zeitschrift für Geschichte und Altertumskunde, vol. 100 (2000), pp. 207–254
Sanz-Esteban I, Calvo-Lobo C, Ríos-Lago M, Álvarez-Linera J, Muñoz-García D, Rodríguez-Sanz D. Mapping the human brain during a specific Vojta’s tactile input: the ipsilateral putamen’s role. Medicine (Baltimore). 2018;97(13):e0253. doi:10.1097/MD.0000000000010253</p>
Shumway-Cook Anne, H. Woollacott Marjorie. (1995). Control Motor. Williams & Wilkins.
Takeshita H, Tanaka, M, Matsuzawa T. Development of Postural Reactions in Infant Japanese Monkeys. The Annual of animal Psichology, 33 (1984)
Vaclav Vojta, Annegret Peters. El principio Vojta. 1992. Springer Verlag-Iberica.
Vojta V, Schweizer E. El descubrimiento de la motricidad ideal. 2011. Ediciones Morata.
«Nada tiene sentido en fisioterapia si no es a la luz de la evolución»
Dobzhansky, genetista y padre de la síntesis evolutiva. En su frase original se ha sustituido «biología» por «fisioterapia».
La motricidad fina no es exclusiva humana
En su obra La edad de la empatía, el primatólogo Frans de Vall relata un curioso hecho observado por él mismo, donde un chimpancé adulto se empleaba con sumo cuidado para sacar una espina que una hembra tenía alojada en su ojo. Y lo consiguió sin causar daño. Un claro ejemplo de motricidad fina de alta precisión.
Otro ejemplo de alta coordinación y precisión manual es el despioje, el acto más social entre los chimpancés. También sabemos que son capaces de elaborar herramientas como ramitas que preparan para introducir en termiteros. Esto revela que la motricidad fina no es exclusiva de nuestra especie, lo que no significa que tengan las mismas habilidades que tiene nuestra mano.
Qué diferencia nuestra motricidad fina?
representación de H.Habilis obteniendo lascas de una piedra
En otro experimento se intentó entrenar a unos chimpancés para obtener filos cortantes de una piedra (lascas). Fue un fracaso. Los investigadores concluyeron que esta actividad requiere de una coordinación y unos movimientos que los chimpancés no tienen disponible: la mano radial.
Mano radial haciendo la pinza de precisión.
La mano radial necesita como «ingredientes» de movimientos en la muñeca la flexión dorsal y desviación radial en combinación con la pronación del antebrazo.
Además, la larga distancia entre su pulgar en relación a los demás dedos resulta determinante para el éxito de esta compleja acción de coordinación.
Por qué no disponen de estos componentes de movimiento?
Locomoción en nudillos
La respuesta esta en su tipo de locomoción cuadrúpeda/arborícola. En otras palabras, no se puede tenerlo todo. Para colgarse de las ramas de los árboles son muy útiles unos largos metacarpianos que facilitan la presa tipo «gancho» junto con unas falanges curvas.
Además, disponer de flexión dorsal de la muñeca daría gran inestabilidad a la articulación de la muñeca al apoyar el peso en su locomoción de nudillos terrestre. Luego lo que nosotros podemos ver como limitaciones a su motricidad fina, para ellos, en su nicho ecológico, son ventajas para su locomoción, puesto que al contrario que nosotros, los chimpancés hacen un uso ambivalente de su mano.
En ese contexto postural cuadrúpedo/arborícola tanto la columna vertebral como la pelvis forman parte de la misma ecuación donde se ubica la cinesiología de la mano. Es decir, los componentes cinesiológicos de la mano son interdependientes del patrón global de la locomoción.
En consecuencia, la posición de la columna vertebral (CV) y la pelvis tendrán una influencia critica en la cinesiología de la mano, y viceversa. Como veremos, la postura axial será de especial importancia en el abordaje de las lesiones neurológicas que interesan a la mano humana y a la postura global.
Entonces, ¿de donde procede nuestra motricidad fina?
Izq: escapula chimpancé. Dcha: humana
La motricidad fina de los chimpancés esta directamente relacionada con el patrón postural global para su locomoción, caracterizado por la cifosis de su CV y la pelvis en anteversión con una glenoides orientadas cranealmente.
De un modo equivalente nuestra motricidad fina esta vinculada con nuestra postura global para la locomoción bípeda, que según el registro fósil, precede a la mano radial (homo habilis) en al menos 2 millones de años. O dicho en otras palabras, nuestra motricidad fina procede de la extensión axial de la CV y la retroversión de la pelvis con unas glenoides escapulares orientadas lateralmente.
El daño neurológico revela nuestra herencia evolutiva.
Observe la postura de la mano
La pérdida de control postural que conlleva una lesión del SNC nos aboca a un estadio postural primitivo en muchos aspectos motores como es el control automático de la postura. Esto es muy evidente en las extremidades del lado afectado y algo menos en el tronco. Pero es precisamente en la CV, la pelvis y las escapulas donde ésta involución postural más afecta a la motricidad de las extremidades.
observe la similitud del patrón con la imagen de arriba
En otras palabras, la CV pierde el control de la extensión axial y aparecen compensaciones en el plano sagital: hipercifosis o hiperlordosis, y en el frontal en forma de excurvación del lado afecto en la zona torácica, provocada por la acción sin oposición de la musculatura epiaxial del lado sano. La cabeza se posiciona asimétricamente fuera de la linea media.
En la pelvis, la consecuencia siempre es la misma, una vuelta a la anteversión primitiva que se manifiesta con un flexo constante en la cadera al caminar. En la escapula del lado afecto se pierde el equilibrio en su función de distribuidor de fuerzas entre la extremidad, la CV y el tronco. Los potentes rotadores internos (pectoral y dorsal ancho) se llevan la articulación, falta congruencia articular y aparecen signos de subluxación.
Entonces, la mano expresa esta situación con tendencia a la flexión palmar, desviación ulnar de muñeca y los dedos flexionados. Un patrón reconocible en los chimpancés.
Postura y función en el abordaje de las secuelas neurologicas en el MS.
Actualmente en el abordaje clínico de las secuelas del ictus en el MS, se concede una importancia más teórica que práctica a la situación postural como contexto para trabajar las habilidades y destrezas funcionales de la mano. Es decir, la prioridad se centra en el MS, mientras que la postura (generalmente en sedestación), es algo secundario. La postura/función comparada con otros primates nos confirma que ambos conceptos están íntimamente interconectados. Esto no se debe interpretar como que el abordaje proximal es prioritario sobre el distal, ni viceversa. Es simultáneo.
De hecho, postura y función trabajan como un conjunto interdependiente durante el neurodesarrollo. Una vez que este proceso es integrado y asimilado en el SNC (entorno a los 4 años), postura y función pueden operar de forma disociada, es decir, podemos manipular con gran precisión con la pelvis en anteversión o con la CV en cifosis.
La postura no pertenece a esa función del MS en la ontogénesis.
Es posible que esta experiencia influya negativamente a la hora de conceder a la postura el mismo grado de importancia que a la movilidad del brazo/mano afecto.
bebe sano, 1º mes
Bajo el paradigma evolutivo las secuelas motoras tras un ictus se interpretan como una disfunción patológica que conlleva en cierta medida una involución (no regresión) a circuitos primarios del SNC de nuestra propia ontogénesis en el primer trimestre.
Hablamos de un estadio de inmadurez postural donde están presentes la anteversión pélvica, déficit de extensión axial de la CV, rotación interna de las articulaciones esféricas (hombro y caderas) y la desviación ulnar de muñecas.
Algunos de estos patrones parciales también podemos reconocerlos en nuestros parientes simios: anteversión pélvica, flexo de cadera y rodilla o la desviación ulnar de la muñeca. Las secuelas tras un ictus no se deben confundir con una degradación a un escalón evolutivo inferior al estilo de la refutada ley biogenética de Haeckel (también conocida como teoría de la recapitulación), sino a la máxima de que se pierde primero lo último que se adquiere en la evolución.
La ontogénesis humana tiene las preguntas… y las respuestas
Desde este punto de vista, la estrategia rehabilitadora se entiende como una vuelta al proceso de neurodesarrollo que implica reiniciar la vinculación del binomio postura-función destruido por la lesión. Teniendo en cuenta que la maduración motora en nuestra especie se produce siguiendo pautas regulares en su interacción con el entrono (Gesell 1952, Vojta 1972), es evidente que este proceso esta guiado por la información genética.
Luego, si la información que contiene las instrucciones de nuestro desarrollo motor esta impresa en el genotipo de cada individuo, desde los elementos más básicos hasta los más complejos como es la motricidad fina de nuestra mano ¿sería posible volver a acceder y activar esta información genética en la edad adulta?
Envía tu nombre y e-mail a: lperales302@gmail.com. y recibirás una invitación (gratis) al website y página de Facebook privada donde se responde a esta pregunta.
La teoría de selección de grupo neural (TSGN) destaca la noción de que el desarrollo motor del bebé es el resultado de un complejo entrelazado de información de genes y medio ambiente(1). La ontogénesis de Vojta esta fundamentada en el programa genético sobre la base del descubrimiento de las Locomociones Reflejas (LR), lo cual no es incompatible con el reconocimiento de las influencias ambientales en el neurodesarrollo, ya que el patrón genético debe interactuar necesariamente con el entorno(2).
Este documento tiene como objetivo resumir las relaciones entre ambas teorías en diez puntos:
1. La variación es una constante durante el neurodesarrollo motor humano(3). La ontogénesis postural (OP) según Vojta identificó, mediante el descubrimiento de las LR(4,5), los patrones posturales y motores parciales universales (genéticos) presentes en el desarrollo normal, tanto en el desarrollo llamado típico como en las variaciones.
2. La OP según Vojta NO describe patrones típicos, sino patrones ideales(6) (genéticos). Se trata de la cinesiología que es coincidente con los patrones típicos correspondientes a la fase de variabilidad primaria de la TSGN, y que subyacen en las variaciones presentes en el desarrollo motor atipico.
3. Por la regularidad en que los patrones ideales (genéticos) se presentan en el desarrollo, la OP según Vojta constituye un instrumento para el reconocimiento de las variaciones patológicas hasta alcanzar la marcha libre (primer año de vida). Las reacciones posturales confirmaron la correlación existente entre OP y la maduración postural del SNC (7-10). Los Movimientos generales de Prechtl, (mencionados en la TSGN) para la detección de alteraciones neurológicas del lactante son igualmente de naturaleza genética.
4. La OP de Vojta se enfoca en la constante genética presente en el desarrollo, lo cual no implica ignorar la interacción con el ambiente que recoge la TSGN. Sin ella, no es posible la integración de los patrones genéticos activados mediante la LR(6).
5. No se puede comprender la OP según Vojta sin conocer los fundamentos de su origen: las locomociones reflejas(11). Patrones globales innatos de origen genético comprobados en estudios neurofisiológicos (12-15).
6. Las variaciones en la OP, como es el caso del gateo, se utilizan para cuestionar la existencia de un programa motor geneticamente predeterminado. Sin embargo, el gateo es la expresión del patrón cruzado (en la locomoción), también presente en otros ítems del desarrollo ideal y típico. Es el patrón cruzado (marcha humana) el que debe estar presente en la OP y no necesariamente en la forma del gateo(2).
7. La variación en la OP no es aleatoria, sino que está determinada por criterios establecidos por la información genética(16,17) plasmada en circuitos neurales pre-establecidos ya en el feto(18). La activación de los patrones parciales de la OP (mediante la LR) permiten un acceso al sustrato genético(6), que puede dar lugar a patrones ideales (típicos para la TSGN), o en su defecto producir variaciones.
8. La pauta genética en el neurodesarrollo motor determina los limites biológicos entre cuyos márgenes puede operar la individualidad como fuente de variación (19), un hecho también recogido en la fase de variabilidad primaria de la TSGN. En esta fase, los modelos predictivos de consecución de la locomoción específica en diferentes especies (incluida la nuestra), indican el claro predominio de la maduración neural sobre el ambiente en TODAS las especies(20)
9. La activación de los programas motores predeterminados mediante la aplicación de las LR es plenamente compatible con la provisión de experiencias sensoriomotoras variables ajustadas al nivel de desempeño del bebe como sugiere la TSGN(1), ya que el entrenamiento potencia los efectos del programa genético, siempre dentro de los limites biológicos.
10. En comparación al neurodesarrollo de otros primates(20), nuestra lenta OP da margen a una mayor influencia ambiental en el despliegue de los programa motores predeterminados, visible en las variaciones que se dan respecto a la OP según Vojta. La TSGN ofrece una explicación coherente a este hecho.
En conclusión, la TSGN y la OP según Vojta lejos de ser incompatibles son claramente complementrias.
Esquema que muestra los patrones posturales ideales (geneticos) por sus relaciones analogas con los patrones parciales presentes en las LR.
Bibliografia
1 Hadders-Algra M. The neuronal group selection theory: a framework to explain variation in normal motor development. Dev Med Child Neurol. 2000;42:566– 72.
2 Vojta V, Schweizer E. El descubrimiento de la motricidad ideal. Ediciones Morata 2011.
3. Touwen BCL. (1993) How normal is variable, or now variable is normal? Early Human Development 34: 1–12
4. Vojta, V. (1970). Reflex rotation as a pathway to human locomotion. Zeitschrift für Orthopädie und Ihre Grenzgebiete, 108(3), 446–452.
5. Vojta, V. (1968). Reflex creeping as an early rehabilitation programme. Zeitschrift für Kinderheilkunde, 104(4), 319–330.
7. Zafeiriou DI. Primitive reflexes and postural reaccions in the neurodevelopmental examination. Pediatr Neuroll 2004; 31:1-8.
8. Zafeiriou DI, Tsikoulas IG, Kremenopoulos GM. Kremenopoulosb, Kontopoulosa EE. Using postural reactions as a screening test to identify high-risk infants for cerebral palsy: a prospective study; 1998:20(5):307-11.
9. Imamura S, Sakuma K, Takahashi T. Follow-up study of children with cerebral coordination disturbance. Brain Dev 1983;5(3):311-4.
10. Zafeiriou DI, Tsikoulas IG, Kremenopoulos GM. Prospective Follow-up of Primitive Reflex Profiles in High-Risk Infants: Clues to an Early Diagnosis of Cerebral Palsy. Pediatr Neurol. 1995 Sep;13(2):148-52.
11. Vojta V. Peters A. El principio Vojta. Springer-Verlag Ibérica 1995
12 Hok P, Opavský J, Kutín M, Tüdös Z, Kaňovský P, Hluštík P. Modulation of the sensorimotor system by sustained manual pressure stimulation. Neuroscience. 21 de 2017;348:11-22.
13 Hok P, Opavský J, Labounek R, Kutín M, Šlachtová M, Tüdös Z, Kaňovský P, Hluštík P. Changes in brain activation after therapeutic stimulation using Vojta therapy: Controlled study. Frente Neurosci . 2019 17 de julio; 13: 722.
14 Gajewska E, Huber J, Kulczyk A, Lipiec J, Sobieska M. An attempt to explain the Vojta therapy mechanism of action using the surface polyelectromyography in healthy subjects: A pilot study. J Bodyw Mov Ther 2018;22(2):287-92.
15. Sanz-Esteban I, Calvo-Lobo C, Ríos-Lago M, Álvarez-Linera J, Muñoz-García D, Rodríguez-Sanz D. Mapping the human brain during a specific Vojta’s tactile input: the ipsilateral putamen’s role. Medicine (Baltimore) 2018;97(13):e0253.
16. Waddington CH. (1962) New Patterns in Genetics and Development. New York: Columbia University Press.
17. Gottlieb G. (1991) Experiential canalization of behavioral development theory. Developmental Psychology 27: 4–13
18. Turk E, van den Heuvel MI, Benders MJ, de Heus R, Franx A, Manning JH, Hect JL, Hernandez-Andrade E, Hassan SS, Romero R, Kahn RS, Thomason ME, van den Heuvel MP. Functional Connectome of the Fetal Brain. J Neurosci. 2019 Dec 4;39(49):9716-9724.
19. Shumway-Cook A, Woollacott MH. Control Motor. Williams & Wilkins; 1995.
20. Garwicz M , Christensson M , Psouni E . A unifying model for timing of walking onset in humans and other mammals. Proceedings of the National Academy of Sciences . 2009;106:21889-21893. https://doi.org/10.1073/pnas.0905777106
La comprensión de la evolución del bipedalismo humano puede proporcionar información valiosa sobre las características biomecánicas, fisiológicas y anatómicas de la locomoción en los humanos modernos, no solo por saber como este proceso aconteció en el pasado, si no por las importantes implicaciones que este conocimiento puede aportarnos en la rehabilitación de la marcha en personas que por diferentes patologías, neurológicas u ortopédicas, se ven limitados o impedidos con considerables costes económicos y sociales. La teoría neuro-plástica surge desde la práctica clínica de la neruorehabilitación con técnicas basadas en cadenas musculares y el principio de locomoción refleja de Vojta en interacción con otras disciplinas como la paloantropologia, biomecánica, neurofisiologia, neurodesarrollo postural, y anatomía comparada. El resultado es una teoría que afronta los «cómos» en lugar de los «porqués«. Que explica como el patrón neural bípedo, disponible en la ontogénesis postural temprana de cualquier primate, fue el responsable del proceso plástico de remodelación anatómica en las crías que adoptaron esta forma de locomoción por la presión selectiva de una comunidad inmersa en el comportamiento bípedo, que no bípeda. Un proceso que dejó marcas epigenéticas indelebles que vincularon al patrón neural bípedo con las estructuras anatómicas que provocó. El análisis de las respuestas posturales y locomotoras reflejas que surgen de la estimulación de estas zonas nos aportan pruebas a la altura del desafío que afronta.
Entre los primeros cambios anatómicos que iniciaron el proceso de transformación, desde una locomoción arborea a otra bípeda existe un amplio consenso, basado en el registro fósil, que tuvo su origen en la pelvis (Coppens 1991). De hecho, esta estructura resulta esencial para determinar con seguridad el bipedismo de un determinado espécimen fósil.
En este sentido el antropologo físico Sherwood Washburn apuntó a que el tránsito desde una pelvis arboríca a otra bípeda fue iniciado por la flexión de los huesos iliacos hacia delante, argumentando que la nueva orientación espacial de estos huesos supuso la reordenación de los vectores de fuerza de los músculos glúteos (Washburn 1968). Con esta acción pasaron de ejercer su acción como extensores de la cadera a ser abductores sin apoyo en el pie.
Sin embargo, su acción más decisiva fue la que realizaron (y realizan) en cadena cerrada, es decir, cuando nos quedamos apoyados sobre una pierna al caminar. Entonces, el glúteo medio con su orientación lateral se encarga de enderezar la pelvis en el plano frontal evitando que se incline hacia el otro lado (Arsuaga y Martinez 1998).
¿Como resuelven este problema los chimpancés la caminar? inclinan el tronco sobre la pierna de apoyo, de forma que el centro de gravedad cae sobre la base de sustentación.
En este análisis, la inclinación lateral del tronco sobre la pierna de apoyo al caminar erguidos de otros primates no humanos se considera como la consecuencia lógica de la orientación posterior de los huesos iliacos (Arsuaga y Martinez 1998). Si a ello unimos que el registro fósil muestra una clara progresión en la flexión de los ilicos, todo parece indicar que Washburn apuntaba en la dirección correcta.
Sin embargo, nunca se ha considerado la posibilidad de que la flexión de los huesos iliacos en dirección ventral fuese consecuencia, y no causa, de otros procesos menos evidentes acontecidos en otra zona del cuerpo.
2. Hipótesis neuro-plástica, una explicación alternativa al surgimiento de la bipedia
Planteamos la teoría de que la bipedestación obligada surgió en el seno de una comunidad que por las razones que fuesen (acarreo, moda, deforestación etc) adoptaron masivamente la locomoción bípeda como forma de desplazamiento habitual arborícora y terrestre, a pesar de no disponer de una anatomía preparada para la bipedia. Las crías de estos ancestros asimilaron de forma precoz el patrón neural de la locomoción bípeda, disponible en la ontogénesis de todo primate, por el potente estímulo de la imitación procedente de toda una comunidad inmersa en el bipedismo.
La locomoción bípeda es habitual entre los Bonobos
De esta forma, un patrón «auxiliar» como es la bipedia para los primates, pasaría a tener un papel principal, modelando la plástica estructura osea de los infantes para imprimir en ella la impronta física del patrón de coordinación bípedo. La pelvis fue en apariencia, la diana principal de estos primeros cambios que, sin embargo, tuvieron lugar en una zona nuclear para la motricidad de cualquier vertebrado: La columna vertebral (CV).
Se argumentará que la orientación iliaca dorsal, propia de los primates no humanos, forma parte de un contexto anatómico-funcional configurado para la locomoción arboricora/cuadrúpeda, en el cual la musculatura autóctona que controla la función de la CV juega un papel esencial. Sostenemos que el cuerpo, en su diseño anatómico-funcional, es un reflejo del patrón neural de locomoción que esta configurado en el genotipo de esa especie. Por tanto, el cambio del patrón neural de locomoción lleva implícito el cambio del diseño anatómico del que se sirve para ejecutarlo. Un proceso de esta naturaleza dejó una enorme cantidad de pruebas en el camino… como veremos más adelante.
3. El proceso hacia la anatomía bípeda.
3.1 La musculatura profunda de la CV, la clave del cambio
Tomado de O´Neill: Muestra el % de fibras Tipo I en chimpancés: 32% por el 69% en humanos
La adopción del comportamiento bípedo de forma «viral» por las crías imitando la pauta social por medio de la neuronas espejo (Rizzolatti 2008) provocó en primer lugar un cambio en la proporción del tipo de fibras musculares que son dominantes en los primates no humanos como el chimpancé, las del tipo II, rápidas y fatigables hacia el tipo I, lentas y resistentes (Oneill 2017).
Esta transferencia afectó a grandes grupos musculares especialmente aquellos que pasaron a tener funciones de sostén del eje axial contra la gravedad como son los que rodean a la CV (Johsom 1973, Regev 2010) gracias a la plasticidad del tejido muscular en función del tipo de actividad a la que son expuestos, sobre todo a edades tempranas (Pette 2000, Staron1997, Wilson2012).
En esta linea, un trabajo de Shapiro (1988) estudió mediante EMG tres músculos de la espalda profunda (multifidus, longissimus thoracis, iliocostalis lumborum) en chimpancés y gibones durante la locomoción erguida, concluyendo que los patrones de actividad de estos músculos son coincidentes con los humanos. Lo que sugiriere que la diferencia se encuentra en la arquitectura de estos músculos, es decir, en la composición de sus fibras.
Tomado de Regev 2010: Porcentaje de fibras tipo I en la muculatura profunda
Musculatura autóctona de la CV
En la región torácica de la CV, esta transición hacia la fibras del tipo I produjo efectos anatómicos y funcionales decisivos (Plomp 2019) que dotaron a este segmento de una gran estabilidad, de modo que las crías de estos antropomorfos podían sostener la CV en su eje axial sin apoyo de los miembros superiores, sin esfuerzo, y por tiempos indefinidos en comparación a los pioneros en el comportamiento bípedo de su comunidad. Un cambio que tuvo que producirse en un plazo relativamente corto (inferior a 5 años)
Esta nueva estabilidad del eje axial alcanzó su punto crítico cuando la musculatura que se relaciona de forma directa e indirecta con la CV encontró aquí un punto de anclaje fijo que nunca antes habían tenido en posición vertical. Entonces, la CV de estos antropomorfos pasó de tener una función enderezadora, propia de los primates cuadrúpedos gestionada por fibras del tipo II, a otra nueva sin precedentes: la antigravitatoria, altamente resistente a la fatiga gracias a la nueva proporción favorable a la fibras tipo I, más resistentes (Shahidi 2017).
La extensión del esqueleto axial provocada por el nuevo rol de la musculatura autóctona se expresó sobre la morfología vertebral elevando el nivel de transición entre la región torácica y la lumbar a la vertebra T11 (Ward 2017), siendo con ello determinante en la morfología de la pelvis (Middleton 2017).
Un cambio revolucionario. De una magnitud similar a desplazar un pilar central de carga en la catedral de San Pedro de Roma. Algo que supondría una reestructuración arquitectónica que haría irreconocible la forma clásica en forma de cruz de su planta. Además, la nueva distribución de fuerzas afectaría a toda la estructura del conjunto. Llevando este símil a nuestra biología podremos entender la reorganización subsiguiente de las fuerzas miofasciales a nivel global.
3.2 Cadenas musculares & patrones de coordinación innatos
¿Como pudo este cambio en la musculatura autóctona de la CV ser responsable de la flexión de los huesos Iliacos que apuntaba Washburn? Para entenderlo es necesario comprender que nuestro aparato locomotor, es mucho más que un conjunto ordenado de huesos, músculos y nervios. Se trata de un intrincado sistema miofascial (músculo y su tejido conectivo) que se encentra organizado en cadenas musculares (Busquet 1996 y Campignion 2002). Las cadenas musculares representan circuitos en continuidad de dirección y de planos, interconectados entre sí, a través de los cuales se propagan las fuerzas organizadoras del cuerpo, y cuya existencia y funcionalidad ha sido demostrada científicamente (Huijing 2001, Krause 2016, Wilke 2016).
En cambio, los patrones de coordinación innatos, como es la marcha, el gateo, o los alcances, representan una compleja combinación en el uso de estas cadenas musculares ejecutadas a través del SNC en una secuencia concreta predeterminada geneticamente (Takakusaki 2017, Minassian 2017). Se trata de un sistema relativamente abierto y flexible, ya que la interacción con el medio nos brinda la posibilidad de incorporar nuevos patrones a través del aprendizaje, desde los más simples como chascar los dedos, hasta los más complejos como hacer el pino, patinar o bailar claque (Schmidt 2018). Sin olvidar, que de forma consciente, también podemos seleccionar movimientos aislados de músculos concretos como por ejemplo mover una falange de un dedo o levantar una ceja.
Este lenguaje de cadenas musculares y patrones de movimiento puede resultar confuso para el lector no familiarizado con esta nomenclatura. Para ayudar a su comprensión piense en las cadenas musculares como las reglas de movimiento que tienen cada una de las figuras del ajedrez, que son seís (peon, Torre, caballo, alfil, reina y rey) y en los patrones como en las combinaciones de jugadas que podemos hacer con las figuras, muchas de las cuales, las más básicas, ya vienen programadas de serie (como en los juegos electrónicos). Muchas jugadas se aprenderán en función de las sucesivas partidas ampliando cada vez más el repertorio y estrategias combinativas entre las piezas.
Así como las reglas de movimiento de las figuras del ajedrez nos dan un marco amplismo para desarrollar las jugadas, la organización y distribución de nuestras cadenas musculares nos brindan el escenario biomecánico en el que actúan los patrones motores con incalculables posibilidades combinatorias. ¿Cómo podría cambiar el juego? cambiando las reglas, bien por un error o ignorancia de los jugadores, que modifique el movimiento de alguna pieza (mutaciones), como por ejemplo que la torre también se mueva en diagonal, o por un cambio en el comportamiento de los jugadores que introduzcan una nueva regla (función).
En cualquier caso al verse afectadas las reglas (cadenas musculares) afectará a las estrategias y combinaciones de juego (patrones) que deben adaptase a las nuevas normas. Entonces se convierte en otro juego. Recuerden, como anécdota metafórica, que el rugby surgió del fútbol, porque de forma inesperada, en contra de las reglas, un estudiante ingles (William Webb Ellis) cogió el balón con las manos y corrió con él hasta la portería sorteando rivales. Ese simple gesto provocó una escisión hacia otro juego distinto.
3.3 La musculatura abdominal, responsable último de la flexión de los iliacos.
La mayoría de los patrones globales son innatos y específicos de cada especie como la locomoción, y otros, fruto del aprendizaje. Estos patrones son la expresión de un complejo circuito neural que implica e interconecta amplias zonas del sistema nervioso central en constante interacción con los estímulos procedentes del entorno.
Cadena espiral
Para el caso que nos ocupa, (la modificación anatómica que impuso la bipedia) de las seís cadenas musculares constatadas (Busquet 1996), abordaremos las cadenas espirales, que describen una especie de «X» y discurren en su tramo troncal desde la CV torácica hasta el hueso iliaco del lado opuesto, cruzándose con la otra cadena diagonal en el abdomen (linea alba). Además, esta cadena incluye a la musculatura espinal de la CV. (imagen izq)
Cadenas musculares que compartimos con otros primates como los chimpancés, pero su actividad es diferente, ya que sus puntos de anclaje siendo anatómicamente similares no operan de la misma manera. La diferencia principal está en el comportamiento de su inserción en la CV torácica durante la locomoción bípeda: de punto fijo inestable en los primates no humanos, a punto fijo estable en los humanos. ( ver anexo 1: la paradoja de Thompsom).
Este punto fijo estable, adquirido en el extremo craneal de la cadena espiral (CV torácica) produjo un cambio en las reglas del juego, provocando una reorganización crítica en los vectores de fuerza implicados en estas cadenas diagonales, ya que durante la locomoción bípeda, en su momento más complejo (cuando estamos apoyados sobre una sola pierna), la mitad superior de la cadena tira hacia su inserción en la columna torácica, mientras que la mitad inferior tira hacia su inserción caudal en el iliaco, que se corresponde con la pierna que apoya en el otro lado.
El centro como punto fijo
¿Qué sucede cuando desde ambos extremos de una misma cadena se tiran hacia sí con fuerzas equivalentes? que en el centro de cadena se concentran fuerzas trabajando con vectores opuestos, lo cual produce un efecto físico paradójico, ya que el centro, al no desplazarse, se constituye en punto fijo para los extremos de la cadena, invirtiendo la dirección de los vectores de fuerza hacia este punto (linea alba).
De este modo, surgieron las fuerzas que traccionaron de las inserciones costales en dirección caudal/medial (linea alba) y con fuerza similar del hueso iliaco con vectores craneal/medial, también en dirección a la linea alba. En este último caso, con predominio de un vector más horizontal por la orientación de las fibras musculares en la región abdominal inferior (transverso y ambos oblicuos). Lo que explica que la deformación del iliaco siguiese una trayectoria medial, también, porque el vector craneal queda compensado con el vector caudal que ejercen el tensor de la fascia lata y el sartorio, ambos con origen en la EIAS. El recto anterior del abdomen no forma parte de esta cadena, (más adelante trataremos su papel en otro plano).
En consecuencia, en el transcurso de no más de cuatro a cinco años bajo el efecto remodelador de este nuevo juego de fuerzas, ambos huesos Iliacos fueron traccionados hacia dentro, concentrando la resultante de fuerzas sobre el iliaco en el angulo superior lateral, formando la gruesa EIAS que conocemos hoy día en el iliaco humano. Los ángulos costales sufrieron el mismo proceso de modelaje con tendencia al cierre de los arcos costales, aunque de forma más progresiva al conservar éstos primeros bípedos cierta vida arborícora.
EIAS
Con austrolopitecus la remodelación de los iliacos ya respondían a las demandas estructurales básicas del patrón neural bípedo, pero la co-existencia con patrones locomotores relacionados con la trepa limitaba la expresión física del patrón bípedo en toda su dimensión. Más tarde, con el abandono total de la vida arborícora, el patrón neural bípedo terminó de completar su trabajo de modelaje sin competencia con otros patrones que implicaban apoyos sobre las extremidades superiores.
Entonces, las cadenas espirales terminaron por «abrochar» los ángulos costales hacia su punto fijo en la linea alba abdominal, confiriendo al tórax su característica forma de tonel. Una vez alcanzado el punto de equilibrio entre las diferentes fuerzas de todas las cadenas implicadas en el tronco, el proceso plástico de modelaje en el tronco dejo de actuar (Homo Erectus).
Tórax en tonel (humano) y campana (chimpancé)
Para entonces, el patrón neural de la locomoción bípeda, que se había iniciado con una modificación en la composición de las fibras musculares encargadas del sostenimiento dinámico de la columna vertebral, se había «trabajado» por imposición de la función una estructura troncal acorde a la biomecanica bípeda.
En ello no es posible atribuir objetivo o intencionalidad alguna al proceso, puesto que las crías (las auténticas protagonistas de esta revolución plástica) no tuvieron opción al adoptar el patrón bípedo. Fueron «víctimas» de una presión selectiva ambiental impuesta desde el vínculo afectivo-emocional (su propia madre) y el de toda una comunidad inmersa en el comportamiento bípedo, que no bípeda.
¿Qué otra opción tenían estos infantes que se convirtieron en los primeros bípedos auténticos? disponían del patrón en su ontogenesis temprana y estaban sometidos al potente estimulo del comportamiento bípedo. Un espejo en el que no podían dejar de mirarse. Plausibilidad biológica y presión selectiva en una única dirección ¿Acaso tiene alguna elección un bebe chino, de padres y entorno social chinos, que vive en china, de no aprender chino?
4. La hipótesis neuro-plástica en las teorías de la evolución
4.1 Teoría neuro-plástica y lamarkismo
Al poner la función como piedra angular del bipedismo en esta hipótesis es inevitable observar cierta relación con la vieja teoría Lamarkista de la herencia de los caracteres adquiridos, pero no es así. Mientras que Lamark entendía que la adopción de una determinada conducta o función adaptativa por un organismo a lo largo de su vida podía transmitirse a su descendencia (recuerden el clásico ejemplo de las jirafas) en la teoría neuro-plástica no hay herencia adquirida de adulto a niño en su génesis.
Niño salvaje desplazándose de forma cuadrúpeda
El proceso opera a la inversa, ya que es la descendencia la que adopta una función postural disponible en su ontogenesis temprana por la presión selectiva impuesta por su grupo social. Algo similar a las experiencias reportadas de los niños, que siendo muy pequeños fueron criados por cuadrúpedos: monos y lobos principalmente (Hurton 1940).
Estos niños adoptaron la locomoción cuadrúpeda sobre manos y pies por dos razones: (1º) porque ese patrón estaba disponible en nuestra ontogenesis temprana, siendo ejecutado con gran facilidad debido a un índice intermenbral favorable a esta locomoción en los infantes hasta los 5 años, donde las extremidades son proporcionalmente más pequeñas que el tronco, y (2º) por la fuerte presión selectiva en esa dirección ejercida por el vínculo afectivo que le une a su grupo social de adopción.
En estos casos tan excepcionales la pregunta que podría hacerse es si los bebés criados en dichos ambientes cuadrúpedos alcanzarían a producir modificaciones estructurales que mejorasen el rendimiento funcional cuadrúpedo por el uso de una función postural y locomotora para la cual su fenotipo no estaba predeterminado. De hecho, las experiencias reportadas tienen en común las dificultades para realizar la marcha bípeda (Hurton 1940). En animales, un ejemplo de esto fue descrito por el veterinario neerlandes E.J. Slijper en una cabra que había nacido con una parálisis de sus patas anteriores, y que se las había ingeniado para caminar con sus patas posteriores.
Cuando la cabra murió, Slijper la diseccionó y descubrió que su estructura anatómica era distinta: había desarrollado nuevos tendones en sus patas traseras y su tren superior también era diferente. Para la anatomista Alice Roberts (2018) estos cambios fueron inducidos por el uso de sus patas traseras, aduciendo que una maleabilidad anatómica de este calado solo sería posible en las primeras etapas del desarrollo.
4.2 Asimilación genética
Y yendo más lejos aún, toda vez, que la selección natural (SN) favorece estos cambios fenotípicos inducidos por la función ¿sería posible incorporados al genotipo para tenerlos disponibles en las generaciones sucesivas? la respuesta a esta cuestión la dió el genetista británico Conrad Waddington a mediados del S-XX, bajo el término «Asimilación genética», un proceso mediante el cual una respuesta fenotípica a un factor ambiental es asimilada por el genotipo de modo que se vuelve independiente del inductor ambiental (Waddington 1951).
Algo que ilustraba con el callo del avestruz. Un claro ejemplo de como una característica que procede de la experiencia ha sido incorporada al genotipo. Waddington explica la adquisición evolutiva de los callos a partir de la asimilación genética, invocando una transferencia de competencia de un inductor externo a uno interno: la piel del avestruz habría adquirido, por selección natural, la capacidad (genética) de desarrollar callos en respuesta a la fricción. En un segundo paso, la piel se habría vuelto capaz de formar callos por inducción embriológica; inductor interno (Gilbert 1991). El mismo proceso podría llevarse a nuestra pelvis, es decir, nuestros tejidos óseos y conectivos habrían adquirido por SN la capacidad de modelarse en respuesta a fuerzas de tracción/compresión inducidas por el uso de un patrón neural de locomoción alternativo. En un 2º paso los tejidos serian capaces de replicar las nuevas estructuras por inducción embriológica.
La bióloga Mary Jane West-Eberhard (2003) sostiene que los cambios anatómicos resultado de nuevos comportamientos también pueden producir variación siendo posible que hereden parte de esos cambios a causa de modificaciones epigenéticas. En comparación con la mutaciones genéticas que producen variaciones al azar, la adaptación fenotípica genera una variación que desde el comienzo facilitan al organismo su adaptación al medio (West-Eberhard 2005).
Para la antomista Roberts, «es muy probable que los individuos con nuevos rasgos anatómicos que se han desarrollado como respuesta a un cierto comportamiento tengan una ventaja en la supervivencia y la reproducción, de modo que el potencial para generar tales cambios se seleccione favorablemente» A finales de siglo XIX el psicólogo americano James Baldwin acuño un idea similar referida sobre todo a la transmisión de comportamientos que incrementaban la supervivencia contra depredadores, bajo el nombre conocido como efecto Baldwin.
Por todo ello la hipótesis neuro-plástica no se puede relacionar con la herencia de los caracteres adquiridos. Más bien se podría hablar de auto-herencia para convertir un patrón auxiliar o secundario (disponible en la OP) en un patrón primario, inducidos por las presiones selectivas del entorno social y afectivo (inductor ambiental). El uso del patrón bípedo reconvertido en prioritario sería la fuente generadora de variación fenotípica que permitió al patrón neural operar de forma más eficiente como sugiere un trabajo de Druelle (2017).
4.3 Teroría neuro-plástica y neodarwinismo
La teoría neodarwinista defiende que una vez aparecida una presión selectiva en una dirección (la bipedia) se verían favorecidas todas las mutaciones que fueran en esa linea en un proceso lento de selección natural de las mutaciones más aptas a tal fin. Por tanto, la bípedia estaría necesariamente ya presente en la comunidad como vector de presión selectiva previa a la aparición de las primeras mutaciones efectivas para poder orientar éstas en la dirección marcada.
Así, la aparición por azar de una ligera flexión en los iliacos en varios individuos a la vez de una misma comunidad, (para así poder reproducirse entre ellos transmitiendo sus variaciones) sería el factor desencadenante que iniciaría el proceso apuntado por Washburn. En otras palabras, primero la estructura y luego la función, un planteamiento condicionado por la comparecencia en la escena científica de uno solo de los actores: la estructura, que es la que deja huella fósil, ¿y la función?
Ardipitecus, hominido arboricora de pelvis bípeda
Desde el punto de vista neural uno de los conflictos de este planteamiento estaría en el caso de que la bipedia no fuese el patrón locomotor hegemónico, como apunta Tim White y el descubrimiento de ardipitecus (Lovejoy 2010) , que sugiere que la bipedia apareció en un entorno boscoso donde la bipedia, la trepa, la braquiación y la cuadrupedia serían los patrones de locomoción utilizados en este entorno (Wolde 2009). Los primates no humanos utilizan estos patrones de forma indistinta, lo cual sugiere que la pelvis simiesca es el diseño que la SN ha favorecido para este uso ambivalente, no la pelvis bípeda.
En este contexto ¿qué probabilidades de éxito tendría esta primera mutación en ese ambiente antes de que la SN hiciese su trabajo? Otra cuestión sería que el patrón bípedo fuese plenamente hegemónico. En ese escenario las mutaciones «correctas» encontrarían el caldo de cultivo perfecto para prosperar. Eso sí, esa lotería tendría que tocar varias veces de forma simultanea en el seno de una comunidad reducida de miembros estimada en no más de 150 ejemplares ¿probabilidades?
La neurología, otro problema a la hipótesis darwinista
La neurología sugiere que en este contexto ecológico una leve flexión de los iliacos producida por mutación al azar, que entrase en conflicto funcional con otros patrones utilizados en el medio arbóreo, ni siquiera habría tenido la oportunidad de «debutar» para influir en la función postural y locomotora antes de que el SNC hubiese corregido o ignorado esa «anomalía» que se desvía del engranaje anatómico-funcional que utiliza el SNC para sus propósitos locomotores.
Deformidad estructural progresiva causada por una función alterada.
La evidencia científica hace tiempo que viene demostrando la capacidad del patrón neural de modelar la plástica estructura osea en cortisimos espacios de tiempo. Lo vemos en el progreso de la deformidad estructural que subyace a la presencia de un patrón alterado por una lesión neurológica como es la parálisis cerebral. Una deformidad que se inicia a partir de una estructura sana y que desde las primeras semanas de vida va incrementándose y consolidando año tras año (Park 2014, Hasler 2020). Algo que no es exclusivo de la parálisis cerebral. Otros cuadros neurológicos adquiridos en adultos como el Ictus, las lesiones medulares y la ELA cursan del mismo modo.
En la situación inversa, donde la anomalía, en lugar de estar en la función neural (como en la paralisis cerebral o el Ictus) se encuentra en la estructura al nacimiento (por mutaciones, tóxicos, enfermedades etc) y que ésta sea de tal calibre que impida de forma grave la ejecución del programa locomotor geneticamente predeterminnado, vemos que, sin cirugía, no existe alternativa motora favorable en el mundo real, (la ciencia ficción es otro asunto)
Ejemplo de deformidad congénita que impide la función predeterminada
Por tanto, en referencia a la supuesta mutación al azar responsable de la flexión iliaca estamos ante un callejón sin salida. Si fue moderada, hubiese sido ignorada o depurada hacia la norma. Si fue excesiva haciendo con ello inviable su reconducción hacia la pelvis simiesca, lo que tendríamos, no es una vía hacia otro patrón de locomoción, sino una grave discapacidad con un pronóstico vital catastrófico a corto plazo.
Para ilustrar esta situación permitanme este símil: imaginen un rebaño de 1000 ovejas gestionado por un experimentado pastor con 4 perros, en el cual, una oveja de forma accidental se escabulle del rebaño siguiendo una senda cuesta abajo que favorece y facilita sus pasos ¿piensa que esta oveja despistada tendría alguna oportunidad de modificar o tan siquiera influir en la dirección del rebaño que dirige el pastor junto a sus perros?
Tenga seguro que en 1000 de cada 1000 veces que esto suceda, antes de que la oveja haya caminado al margen del rebaño más de 20 metros ya estaría de nuevo en el redil. Es el pastor quien tiene la capacidad de dirigir o modificar la dirección de su rebaño con ayuda de sus perros. Solo en caso de que un terremoto fracturase el terreno y se tragase al rebaño, éstas habrían cogido accidentalmente un camino diferente… un camino hacia ninguna parte. O tal vez, quedase dividido con unas cuantas ovejas aisladas al otro lado de la sima, ¿alguna posibilidad de supervivencia?
En este símil, como seguramente ya se habrá percatado, la senda que toma la oveja representa la presión selectiva, el pastor el patrón de locomoción predeterminado geneticamente en el SNC, los perros el SNC subcortical (acciones automáticas), y las ovejas el sistema anatómico-funcional (operadores del SNC)
Con esto no se pretende discutir la teoría neodarwinista en su conjunto, sino señalar sus debilidades en esta materia (el bipedismo), ya que en cualquier caso los cambios anatómicos producidos, como resultado de la implementación del patrón neural bípedo en las crías, (siguiendo la presión selectiva generada en su comunidad) tuvieron que someterse necesariamente al escrutinio de la SN.
La teoría neuroplástica partiendo de la misma presión selectiva hacia la bipedia que comparte con la teoría darwinista, apunta al cambio en la función como generador de los cambios en la estructura, (y no a la inversa) algo que supuestamente no es posible demostrar, ya que la función, al contrario que la estructura no dejó huella fósil ¿o tal vez sí?
La función no solo dejó huellas fósiles reconocibles en los huesos, si no que al hacerlo también lo hizo en el genotipo, donde estructura y función quedaron vinculadas como la última actualización «hardware/software» para la locomoción. A continuación analizaremos algunas de estas huellas de la función del patrón neural bípedo en la pelvis. Se trata de pruebas que ponen en cuestión las mutaciones al azar como origen de estos relieves, ya que de ser así, su estimulación no debería estar vinculada a un patrón cinesiológico relacionado con la marcha bípeda.
No debería…pero sucede.
5. Las Zonas de activación de La Locomoción refleja de Vojta: los «fósiles actualizados» de la función
Entre 1954 y 1972 el neuropediatra Checo Vaclav Vojta descubrió de forma empírica en personas con daño cerebral congénito que presionando determinadas zonas del cuerpo en posturas concretas en decúbito (supino, lateral y prono), el SNC respondía de forma refleja activando patrones de locomoción global cuyos contenidos cinesiológicos parciales podían ser reconocidos en distintos periodos del neurodesarrollo del niño desde el nacimiento hasta la marcha (Vojta 1972). Periodo conocido como ontogenesis postural o del enderezamiento.
Posición de partida y localización de las zonas de activación en el patron de la Reptación refleja
Al conjunto de sus investigaciones y descubrimientos los denominó Locomoción refleja. Locomoción, porque los tres patrones innatos descubiertos reunían las características de toda locomoción descritas por Grillner (1981) y refleja porque su activación se producía sin la participación voluntaria o consciente por parte del sujeto. Vojta demostró que su activación se producía a cualquier edad, pero con mayor intensidad en los lactantes con efectos muy positivos para su desarrollo (Vojta 1991).
Vojta aplicando su terapia a un bebé prematuro
En la actualidad su empleo terapéutico en hospitales y centros de rehabilitación especializados de todo el mundo permite estimular patrones del desarrollo normal no solo en bebés con riesgo de desarrollar secuelas motoras tras un daño neurológico sino también en adultos (Laufens 1999, Kanda 2004, Senthilkumar 2011, Kotnik 2012, Epple 2020).
La evidencia científica ha demostrado en estudios de nivel 1 empleando RMf (Hok 2017, Hok 2019, Sanz 2018) la singularidad de la estimulación de estas zonas anatómicas en el SNC. Lo mismo han reflejado estudios empleando EMG (Pavlu 2000, Gajewska 2018), en la evocación de los patrones cuadrúpedos de la LR íntimamente relacionados neurofisiologicamente con la marcha bípeda (Diezt 2001, Zehr 2004) .
Sin embargo, hasta la fecha se desconocen su origen y significación, y por qué, cuando alguna de estas zonas reciben una presión determinada con direcciones vectoriales concretas, el SNC responde activando patrones que pertenecen al proceso de neurodesarrollo que incluye la locomoción bípeda.
5.1 Patrones atávicos & patrones modernos
reflejo primitivo de agarre en un neonato
Todos los componentes cinesiológicos de los patrones evocados de forma refleja mediante la estimulación de las zonas descubiertas por Vojta pertenecen a nuestra motricidad actual (Vojta 2011). Con la estimulación de las zonas de la locomoción refleja no tenemos acceso a patrones posturales o motores extintos o atávicos, aunque parte de éstos sí podemos verlos en la patología neurológica y en algunos reflejos primitivos del neonato (Rao 1999). Desde nuestro actual sistema solo tenemos acceso a la última versión de nuestro desarrollo locomotor.
Para hacerse una idea recurriré a la clásica metáfora informática. Si usted adquiere un ordenador que viene con la ultima versión de Windons 10 instalada, no podrá operar con elementos de versiones anteriores aunque la última versión este «armada» sobre la base de todas las precedentes. En otras palabras, desde la versión postural y locomotora Sapiens solo tenemos acceso a patrones motores de esta última versión, que en lo postural y motor, tal vez sea la misma desde Homo erectus (Vojta 1972).
5.2 El significado de las zonas de activación de Vojta: un legado de la evolución
La teoría neuro-plástica justifica que el origen de las zonas descritas por Vojta, junto a otras investigadas (Perales 2013) surgieron en diferentes etapas de nuestra evolución como marcas indelebles de un proceso neuro-plástico guiado por un cambio en el comportamiento postural y motor. Del mismo modo que el cráter de un volcán esta indisolublemente relacionado con las fuerzas telúricas que lo hicieron emerger, dejando en el terreno evidencias que nos permiten reconstruir el escenario de su actividad, las zonas descritas por Vojta, como tejidos vivos van mucho más allá de esta metáfora, ya que la memoria de su emergencia permanece activa durante toda la vida y su desencadenamiento responde a un sencillo código de presión (sensitivo y propioceptivo) en una dirección vectorial determinada y una postura con una relación intersegmentaria concreta (Vojta 1992).
Sostenemos en base a la evidencia, que factores epigenéticos (Legoff 2019, Sherma 2017 Nicoglou 2017) fueron responsables de asimilar en el genotipo de los infantes los patrones neurales responsables de la aparición de estas huellas características en los tejidos que han llegado hasta nosotros, y de las cuales, solo las marcas óseas nos han permitido seguir su rastro a lo largo del registro fósil. Gracias a la vinculación en el genoma del patrón neural junto con las novedades anatómicas que provocaron, disponemos de una puerta de acceso a la evocación de dichos patrones motores con fines terapéuticos. Unas estructuras anatómicas que guardaron recuerdo de este proceso de nuestra evolución, por lo que las denominaremos Zonas de Memoria Evolutiva (ZoME)
5.3 Las ZoME activan patrones no cadenas musculares.
Es importante destacar que la estimulación de las ZoME activan patrones de locomoción innatos, no cadenas musculares. Los patrones utilizan las cadenas musculares como los coches las carreteras. Si consideramos un viaje de Madrid a Chipiona como un patrón, la combinacion de las distintas carreteras utilizadas equivaldría a la combinación de músculos de diferentes cadenas musculares implicadas en la ruta. Si el viaje es muy largo y completo por toda la geografía nacional (patrón de marcha) utilizaríamos casi todas las carreteras importantes en combinaciones muy variables en función de la carretera de salida. Es decir, podemos hacer el mismo viaje desde recorridos diferentes, e incluso, podemos salir desde varios puntos a la vez, imaginado que esto fuera posible, (sumando dos o tres ZoMEs) para completar el viaje más rápido.
Por tanto, la activación del patron de la 1ª posición (ver anexo 2) que inicia en posición fetal y finaliza en bipedestación e inicio de la marcha, desde una ZoME, como por ejemplo la EIAS activa el patrón completo iniciando el proceso desde una ZoME que pertenece a la cadena espiral (la carretera). Si además sumamos otra ZoME, el patrón progresará antes hasta su objetivo.
En este primer trabajo sobre la teoría neuro-plástica nos centraremos principalmente en las ZoMEs que surgieron en relación a la adopción de la postura bípeda en la pelvis.
6. Las pruebas
6.1 ZoME 1:El tubérculo púbico, los rectos abdominales y la retroversión de la pelvis.
Tubérculo púbico hiperdesarrollado por la acción de los rectos abdominales
Empecemos por el final, ¿qué es la retroversión y cómo surgió este movimiento en la pelvis? como se ha descrito, la configuración de un nuevo punto fijo en la región torácica de la CV, no solo afectó a la distribución de fuerzas abdominales pertenecientes a la cadena espiral (transverso y oblicuos), tambíen se vieron involucrados los rectos abdominales pertenecientes a la cadena muscular flexora.
Los rectos, al tener su punto fijo a lo largo de la linea alba, ejercieron una fuerte tracción hacia su centro desde ambas inserciones. En la craneal (esternón y en el borde medial de las últimas costillas) actuando con un vector caudal descendiendo el tórax en su parte central, y en la inserción caudal, traccionando de los tubérculos púbicos en sentido craneal, engrosando su volumen, al tratarse, a diferencia de las costillas, de una estructura menos flexible.
Dicho engrosamiento púbico quedó vinculado al patrón neural causante.
Acción de retroversión (isquios y rectos) y anteversión
Esta acción de los rectos abdominales tuvo una consecuencia aún más importante: la basculación de la pelvis en su conjunto hacia atrás (dorsal), es decir, la retroversión. Una acción donde los rectos actuaron en sinergia con los isquitibiales, que en bipedestación trabajan tirando del isquion hacia sus puntos de inserción caudales.
Así, rectos e Isquiiotibiales fueron los responsables de configurar la nueva posición de la pelvis en el plano sagital. Una posición que redujo completamente la característica flexión del tronco en relación a la cadera propia de los primates no humanos cuando están erguidos, y que fue causante de la aparición de la lordosis lumbar (Kapandji 2006)
Consecuencias de la retroversión para la morfología pélvica:
Disposición de los isquis perpendicular en los simios frente a aparalela en humanos
1ª La reducción del Isquión. Mientras que en la posición cuadrupeda isquión y fémur mantenían una relación próxima al ángulo recto, la posición bípeda sitúa ambos huesos en paralelo acortando considerablemente la longitud relativa de la musculatura isquiotibial. Este hecho eliminó la tensión tendinosa sobre la inserción craneal en el isquion, un estímulo crítico para su crecimiento.
La consecuencia fisiológica fue la reducción de la longitud del isquion hasta encontrar su nuevo punto de tensión (mediado por la aferencia de los órganos tendinosos de Golgi) en relación a la longitud efectiva de unos músculos isquiotibiales reajustados para cumplir de forma eficiente su función.
La retroversión (izq) posiciona el diafragma en paralelo al suelo pélvico
2ª El acortamiento y ensanchamiento de los huesos iliacos, que debido a su innovadora orientación vertical recibieron directamente la presión constante del diafragma en su descenso inspiratorio, junto con las fuerzas procedentes de la musculatura abdominal (activadas concentricamente) y la respuesta opuesta del diafragma pélvico.
La resultante de esta concentración de fuerzas en el Core fue un potente incremento de la presión intra-abdomial que compactó los iliacos liberando en su reducción longitudinal la 5ª vertebra lumbar (encajada entre los iliacos en los primates no humanos). A todo esto hay que sumar el peso de las vísceras abdominales, contribuyendo en conjunto a conformar su característica forma de cuenco.
Consideren que los simios erguidos carecen de retroversión, y por tanto, sus iliacos permanecen orientados hacia ventral en el plano sagital, lo que potencia su acentuada protusión abdominal en posición bípeda.
ZoME del tubérculo púbico (no descrita por Vojta)
La estimulación del tubérculo púbico con vectores craneal/ dorsal/medial activa el patrón global de la Locomoción refleja que determina la posición de partida: supino para el patrón del Volteo reflejo (no descrito en este trabajo), decúbito prono para el patrón de la reptación refleja (no descrito en este trabajo), o posición fetal para el patrón de la 1ª posición (Anexo 2).
El tubérculo púbico pertenece a la cadena flexora por lo que es muy probable que la activación del SNC desde esta zona se inicie a partir de esta cadena, siendo más accesible para su estimulación desde la posición de decúbito supino.
6.2ZoME 2: La Espina Iliaca Antero Superior (EIAS)
La EIAS es un relieve oseo reconocible en todos los primates. Sin embargo, en nuestra especie esta singularmente marcada por su orientación y grosor que permite una fácil palpación desde ventral. Como se ha comentado nuestra particular EIAS surgió por el efecto combinado de fuerzas tractoras en este punto por parte de la cadena espiral (transverso y oblicuos) y el movimiento de retroversión que contribuyó a la generación de la avulsión a causa de la distensión provocada por parte de sus inserciones del sartorio y tensor de la fascia.
ZoME EIAS (descrita por Vojta)
La estimulación de la EIAS con vectores dorsal/medial y resultante hacia la EIPS activa el patrón global de la Locomoción refleja que determina la posición de partida. La EIAS pertenece a la cadena espiral por lo que es muy probable que la activación del SNC desde esta zona se inicie a partir de esta cadena espiral, con especial incidencia, en consecuencia, sobre sobre la musculatura abdominal (transverso y oblicuos) y los movimientos flexores correspondientes a la pierna del lado estimulado.
Dentro del patrón innato de la 1ª posición (ver anexo 2), ya desde la posición fetal de partida, comienzan a activarse componentes intrínsecos y específicos de la marcha humana como son la extensión axial de la columna vertebral, la retroversión de la pelvis y la rotación externa de las articulaciones esféricas, componentes que se mantienen durante toda la dinámica del patrón hacia la bipedestación, y que están plenamente activas en la marcha (Perales 2009, Husarova 2005). A este patrón se accede desde cualquier ZoME individual o en combinación con otras ZoMEs.
Secuencia de activación refleja del patron de la 1ª posición hasta la bipedestación desde la zona EIAS en combinación con zona glutea
6.3 ZoME 3:La Espina Iliaca Antero Iferior (EIAI)
EIAI inexistente en chimpancé
A diferencia de la EIAS, la EIAI no existe en otros primates y su aparición esta datada hace 4 MA coincidiendo con los inicios de la bipedia. De hecho, cuenta con su propio núcleo de osificación.
Su emergencia esta directamente relacionada con el movimiento de retroversión, en un mecanismo de acción idéntico y simultáneo al que contribuyó a formar la EIAS: distensión del músculo recto anterior del cuadriceps en su inserción ósea proximal, generando una nueva avulsión como respuesta a la tracción constante en un tejido tan plástico a una edad temprana.
Según la teoría neuro-plástica, el patrón de triple enderezamiento (tronco, cadera y rodilla) que generó esta avulsión ósea se memorizó en el genotipo junto con la estructura que provocó. Por tanto, ambos eventos (patrón neural y espina) están epigeneticamente vinculados, de modo que al presionar la ZoME con una dirección vectorial dorsal/caudal, el SNC imprime el patrón en su conjunto.
ZoME EIAI (no descrita por Vojta)
La estimulación de la EIAI con vectores caudal/ dorsal/medial activa el patrón global de la locomoción refleja que determina la posición de partida a partir de la cadena extensora a la que pertenece la EIAI, por lo que su estimulación será más efectiva en patrones que impliquen a la cadena extensora de forma segmental (una pierna) o globalmente (todo el cuerpo) como es el caso del patrón de la 1ª posición.
Tomando el ejemplo de este patrón (1ª posición) la estimulación de la EIAI se puede hacer desde la posición de partida, pero es más accesible y efectivo a partir del inicio del enderezamiento del tronco sobre ambos pies que se corresponde con la 6ª posición de este patrón.(Anexo 2).
Estimulación bilateral de la EIAI: Activación refleja del patrón de verticalización humana.
6.4 ZoME 4: La fascia del glúteo medio
Pilar iliaco
Tratándose de tejido conectivo (fascia y musculo) esta ZoME no dejó rastro fósil directo, no obstante, el epicentro de esta zona se encuentra un relieve óseo específico de la bipedia. Se trata del «pilar iliaco» descrito por Arsuaga, una estructura que coincide con el eje de torsión longitudinal por donde los músculos oblicuos flexionaron los huesos iliacos. Sospechamos que el reforzamiento de este eje fue la respuesta fisiológica a los mecanismos de torsión que actuaron sobre el hueso
Zona glútea
Como consecuencia de esta torsión se produjo un alejamiento de los glúteos mayor y mediano, provocando una fuerte distensión en la fascia que conecta ambos músculos. Entonces, fascia, pilar de torsión y patrón neural quedaron epigeneticamente vinculados.
ZoME Glútea (descrita por Vojta)
Su localización a caballo entre dos músculos que pertenecen a cadenas musculares distintas, (glúteo mayor: extensora y glúteo medio: espiral), hacen que su estimulación con vectores ventral/medial actúe con especial incidencia (dentro del patrón de la LR seleccionado) sobre ambas cadenas.
Tomando como ejemplo el patron de la 1ª posición, (anexo 2) que parte de la posición fetal, la ZoME glútea es facilmente accesible desde la posición de partida. El proceso hacia el patrón de la bipedia se inicia, con alta probabilidad, desde la cadena extensora/espiral.
6.5 Otras ZoMEs
El proceso de transformación hasta una anatomía plenamente bípeda comenzó en la parte nuclear de cualquier vertebrado, el eje axial, seguida de la pelvis. Los cambios subsiguientes en las extremidades se dieron como respuesta adaptativa. El patrón neural bípedo no disponía de un plano preconcebido de cuerpo al que dirigirse, tan solo de la función a realizar, ya que nunca antes se había implementado este tipo de locomoción. Por lo tanto, la anatomía tuvo que ir modelándose a través de la función, atrofiando y perdiendo estructuras en desuso, como la abdución del pulgar del pie, y potenciando por la práctica aquellas implicadas en la nueva función, como la configuración del arco plantar.
Estos procesos adaptativos secundarios generaron sus propias modificaciones anatómicas claves en huesos y tejidos blandos. Lugares donde surgieron nuevas ZoMEs vinculadas al patrón neural causante a lo largo de nuestra evolución. Dada su extensión, estas ZoMEs serán objeto de tratamiento en próximos trabajos.
7.Cambio simultaneo o gradual
Una cuestión frecuente en la evolución de la bipedia gira en torno a la dimensión y secuencia temporal que los cambios anatómicos necesitaron para transformar un cuerpo diseñado para locomoción cuadrúpeda/arborícora en otro para la bípeda. ¿Fueron graduales, con cientos o miles de años para que las mutaciones fueran perfilando las bases anatómicas elementales en la pelvis, y que secundariamente dieran lugar a las adaptaciones posteriores en las extremidades? ¿Serían factibles para la SN unos híbridos que pierden especialidades poderosas para quedarse en tierra de nadie largo tiempo hasta alcanzar su nuevo espacio ecológico?
La teoría de «equilibrio puntuado» de Niles Eldredge y Stephen Jay Gould apunta, basado en el registro fósil, a cambios rápidos como respuesta a eventos geológicos y/o climáticos repentinos. La deforestación podría ser uno de esos factores que empujaron a estos homininos a pisar tierra firme, una teoría descartada por el equipo de Tim White y el descubrimiento de Ardipitecus, pero que ha vuelto de nuevo a la actualidad de la mano del Geologo Thure Cerling (2010) que pone en cuestión la teoría de que la bipedia surgió en un entorno plenamente boscoso. En cualquier caso no es objeto de este trabajo entrar en este debate, puesto que nos interesan los «cómos» y no los «porqués».
Sobre el factor temporal, la teoría neuro-plástica plantea que, toda vez que el comportamiento bípedo de esa comunidad (o comunidades independientes) alcanzase una masa crítica suficiente entre sus miembros como para que dicho comportamiento se «viralizase» en la norma social, el proceso, a partir de entonces, apenas se tomaría entre 1 y 3 generaciones en modificar las estructuras nucleares: tronco, pelvis y fémur, siendo plenamente funcionales para operar en tierra firme conservando rasgos arborícoras.
7.1 Progresividad o simultaneidad
Este periodo temporal tan corto también responde a la cuestión sobre progresividad o simultaneidad. Los cambios nucleares se producirían simultáneamente, ya que el patrón neural bípedo lleva implícito operar de forma simultánea en los tres planos del espacio. Una vez configurado el núcleo anatómico-funcional del patrón neural bípedo (tronco y pelvis/caderas) las extremidades progresivamente irían modelando sus estructuras en sincronía con el tronco, lo cual correspondería con un periodo progresivo de afinamiento adaptativo a las necesidades funcionales derivadas de la bipedia como la carrera (Bramble 2004). Otras serían fruto de lo que Gould denominaba exaptaciones, estructuras que en su reorganización en base al cambio anatómico-funcional que supuso al bipedia, dieron lugar casualmente a nuevas posibilidades funcionales como los lanzamientos de precisión y nuevas habilidades manuales.
En resumen, la pelvis cambio de una vez, y de forma simultánea durante los primeros años de vida con el patrón neural bípedo operando a pleno rendimiento en todos los infantes de la comunidad al mismo tiempo. Una idea aceptada y apoyada desde el darwinismo más ortodoxo representada por Ambrosio Garcia Leal (2013):
«Un factor ambiental puede afectar afectar de inmediato al fenotipo de muchos individuos a la vez, mientras que una mutación genética solo afecta inicialmente a un individuo. Los rasgos inducidos por el ambiente son relativamente inmunes a la eliminación por SN en comparación con los inducidos geneticamente…con lo que se incrementa la probabilidad de acomodación genética favorable bajo selección.«
¿Quien imita a quien?
Sin olvidar el enorme impulso de retroalimentación que supone el uso de un patrón bípedo entre los infantes de una misma comunidad. La imitación entre iguales es más poderosa que entre sujetos de diferentes edades (Harris 2015). Es posible que por genética la maleabilidad nero-plástica de los tejidos óseos no fuese la misma en todas las crías, por lo que éstos individuos no alcanzarían a ser buenos bípedos en una sociedad con una fuerte presión selectiva en esa dirección. En pocas generaciones su resistencia plástica al cambio se iría depurando al situarse al margen de la norma, o como diría Dawkins fuera de la moda.
8. Conclusiones
La plasticidad fenotípica junto con la variabilidad de patrones posturales disponibles en la ontogénesis temprana fueron los factores que brindaron la posibilidad de transitar rutas motoras alternativas a las que geneticamente tenían predefinidas nuestros ancestros prebípedos. En ello, el potente estimulo de la imitación del comportamiento bípedo, por parte de unas crías inmersas en bipedismo, supuso el catalizador para la transformación de la costumbre en norma y la norma en biología. Un proceso neuroplástico que dejo huellas indelebles íntimamente vinculadas en el genotipo al patrón neural que las provocó. Zonas que como «notarios» de la evolución guardaron memoria de los procesos neuroanatómicos sucedidos.
El neuropediatra vaclav Vojta descubrió diez de estas zonas de memoria evolutiva junto con las posiciones mas eficaces para su activación con el objetivo de proporcionar en niños y adultos con daños neurológicos patrones ideales de una ontogenesis postural que se remonta al menos hasta Homo erectus. La teoría neuro-plástica brinda una explicación a los orígenes del bipedismo basada en la evidencia científica disponible en biomecánica, neurofisiologia, y anatomía funcional comparada ofreciendo desde una disciplina como es la neurorehabilitación una explicación fundamentada sobre del origen y significación de las zonas descubiertas por Vojta y su íntima conexión con el proceso evolutivo que nos llevó hasta la bipedestación y locomoción bípeda.
9. Bibliografia
Coppens Y. L’originalité anatomique et fonctionnelle de la première bipédie [The anatomical and functional origin of the first bipedalism]. Bull Acad Natl Med. 1991;175(7):977-993. (el bipedalismo empezó edesde la pelvis)
Washburn SL. 1968. The study of human evolution. Eugene, OR: Oregon State System of Higher Education.
Arsuaga JL, Martinez Mendizabal I. La especie elegida. Temas de Hoy. 1998
Rizzolatti G, Sinigaglia C. 2008. Mirrors in the Brain. How We Share our Actions and Emotions. Oxford University Press.
O’Neill MC, Umberger BR, Holowka NB, Larson SG, Reiser PJ. Chimpanzee super strength and human skeletal muscle evolution. Proc Natl Acad Sci U S A. 2017;114(28):7343-7348. doi:10.1073/pnas.1619071114
Johnson MA, Polgar J, Weightman D, Appelton D. Data on the distribution of fibre types in thirty-six human muscles. An autopsy study. J Neurol Sci 1973; 18:111–129.
Regev GJ, Kim CW, Thacker BE, et al. Regional Myosin heavy chain distribution in selected paraspinal muscles. Spine (Phila Pa 1976). 2010;35(13):1265-1270. doi:10.1097/BRS.0b013e3181bfcd98
Pette D, Staron RS. Myosin isoforms, muscle fiber types, and transitions. Microsc Res Tech. 2000;50(6):500-509. doi:10.1002/1097-0029(20000915)50:6<500::AID-JEMT7>3.0.CO;2-7
Wilson JM, Loenneke JP, Jo E, Wilson GJ, Zourdos MC, Kim JS. The effects of endurance, strength, and power training on muscle fiber type shifting. J Strength Cond Res. 2012;26(6):1724-1729. doi:10.1519/JSC.0b013e318234eb6f
Staron RS. Human skeletal muscle fiber types: delineation, development, and distribution. Can J Appl Physiol. 1997;22(4):307-327. doi:10.1139/h97-020
Shapiro LJ, Jungers WL. Back muscle function during bipedal walking in chimpanzee and gibbon: implications for the evolution of human locomotion. Am J Phys Anthropol. 1988;77(2):201-212. doi:10.1002/ajpa.1330770208
Plomp K, Viðarsdóttir US, Dobney K, Weston D, Collard M. Potential adaptations for bipedalism in the thoracic and lumbar vertebrae of Homo sapiens: A 3D comparative analysis. J Hum Evol. 2019;137:102693. doi:10.1016/j.jhevol.2019.102693
Shahidi B, Hubbard JC, Gibbons MC, et al. Lumbar multifidus muscle degenerates in individuals with chronic degenerative lumbar spine pathology. J Orthop Res. 2017;35(12):2700-2706. doi:10.1002/jor.23597
Ward CV, Nalley TK, Spoor F, Tafforeau P, Alemseged Z. Thoracic vertebral count and thoracolumbar transition in Australopithecus afarensis. Proc Natl Acad Sci U S A. 2017;114(23):6000-6004. doi:10.1073/pnas.1702229114
Middleton ER, Winkler ZJ, Hammond AS, Plavcan JM, Ward CV. Determinants of Iliac Blade Orientation in Anthropoid Primates. Anat Rec (Hoboken). 2017;300(5):810-827. doi:10.1002/ar.23557
Busquet L. Les chaînes musculaires. Tome I, Tronc et colonne cervicale. Paris: Frison-Roche; 1996
Campignion P., Denys-Struyf G. Cadenas musculares y articulares concepto GDS. Aspectos biomecánicos. Nociones de base.Alicante: Editorial Such Serra; 2002
Huijing PA,Baan GC. Myofascial force transmission causes interaction between adjacent muscles and connective tissue: from blunt dissection and compartmental fasciotomy in characteristics of the long extensor muscle length of the fingers. Arco Physiol Biochem. 2001; 109 (2): 97 -109.Available from: https://www.ncbi.nlm.nih.gov/pubmed/11780782.14-
Wilke J, Krause F, Vogt L, Banzer W. What Is Evidence-Based About Myofascial Chains: A Systematic Review. GD Arch Phys Med Rehabil. 2016 Mar 1;97(3):454–61. Available from: http://www.ncbi.nlm.nih.gov/pubmed/26281953
Takakusaki K. Functional Neuroanatomy for Posture and Gait Control. J Mov Disord. 2017;10(1):1-17. doi:10.14802/jmd.16062
Minassian K, Hofstoetter US, Dzeladini F, Guertin PA, Ijspeert A. The Human Central Pattern Generator for Locomotion: Does It Exist and Contribute to Walking?. Neuroscientist. 2017;23(6):649-663. doi:10.1177/1073858417699790
Schmidt, R. A., Lee, T. D., Winstein, C., Wulf, G., & Zelaznik, H. N. (2018). Motor control and learning: A behavioral emphasis. Human kinetics.
Hutton, J. H. (1940): «Wolf-children». En: Folklore, transactions of the folk-lore society, vol. 51, n.º 1, págs. 9-31, Londres: William Glaisher Ltd., 1940
Alice Roberts. La increíble probabilidad del ser. Pasado & Presente (2018)
Waddington, C. H., 1941, Canalization of development and the inheritance of acquired characteristics, Nature 150: 563-565.
Gilbert SF. Induction and the origins of developmental genetics. Dev Biol (N Y 1985). 1991;7:181-206. doi:10.1007/978-1-4615-6823-0_9
West-Eberhard MJ. 2003. Developmental plasticity and evolution. New York: Oxford University Press.
West-Eberhard MJ. Developmental plasticity and the origin of species differences. Proc Natl Acad Sci U S A. 2005;102 Suppl 1(Suppl 1):6543-6549. doi:10.1073/pnas.0501844102
Druelle F, Aerts P, Berillon G. The origin of bipedality as the result of a developmental by-product: The case study of the olive baboon (Papio anubis). J Hum Evol. 2017;113:155-161. doi:10.1016/j.jhevol.2017.07.010
Lovejoy CO, Suwa G, Spurlock L, Asfaw B, White TD. La pelvis y el fémur de Ardipithecus ramidus: la aparición de la marcha erguida [la corrección publicada aparece en Science. 12 de febrero de 2010; 327 (5967): 781]. Ciencia . 2009; 326 (5949) :.
Wolde Gabriel G, Ambrose SH, Barboni D, et al. The geological, isotopic, botanical, invertebrate, and lower vertebrate surroundings of Ardipithecus ramidus. Science. 2009;326(5949):. doi:10.1126/science.1175817
Park N, Lee J, Sung KH, Park MS, Koo S. Design and validation of automated femoral bone morphology measurements in cerebral palsy. J Digit Imaging. 2014;27(2):262-269. doi:10.1007/s10278-013-9643-2
Hasler C, Brunner R, Grundshtein A, Ovadia D. Deformidades de la columna en pacientes con parálisis cerebral; el papel de la pelvis. J Child Orthop . 2020; 14 (1): 9-16. doi: 10.1302 / 1863-2548.14.190141
Grillner S (1981) Control of locomotion in bipeds, tetrapods and fish. In: Brookhart JMaMVB (ed) Handbook of physiology. The nervous system. Motor control. American Physilogy Society, New York, pp 1179–1236.
Vaclav Vojta, Annegret Peters. El principio Vojta. 1992. Springer Verlag-Iberica.
G. Laufens, S. Seitz, G. Staenicke, Fachbereich. Fundamentos biológicos comparativos respecto a la locomoción innata, en particular respecto a la “reptación refleja” según Vojta. Fisioterapia 43. Jg (5/1991), Pag. 448-156.
Gajewska E, Huber J, Kulczyk A, Lipiec J, Sobieska M. An attempt to explain the Vojta therapy mechanism of action using the surface polyelectromyography in healthy subjects: A pilot study. J Bodyw Mov Ther. 2018 Apr;22(2):287-292.
Pavlu D, Véle F, HavlíˇckováL . Electromyography and kinesiological analysis of Vojta´s therapeutical principle. Rehab Fyzil. 2000;7:34-9
Kanda T, Yuge M, Yamori Y, Suzuki J, Fukase H. Early physiotherapy in the treatment of spastic diplegia. Dev Med Child Neurol 1984;26(4):438-44. 15.
Ha SY, Sung YH. Effects of Vojta method on trunk stability in healthy individuals. J Exerc Rehabil. 2016; 12: 542-547.
Kotnik SS. Vojta in rehabilitation of children with neurodevelopmental disorders. Paediatr Croat 2012;56(Supplement 1):227-31
Epple, C., Maurer-Burkhard, B., Lichti, M. et al. Vojta therapy improves postural control in very early stroke rehabilitation: a randomised controlled pilot trial. Neurol. Res. Pract.2, 23 (2020). https://doi.org/10.1186/s42466-020-00070-4
Lim H, Kim T. Effects of vojta therapy on gait of children with spastic diplegia. J Phys Ther Sci. 2013; 25: 1605-1608.
Kanda T, Pidcock FS, Hayakawa K, Yamori Y, Shikata Y. Motor outcome differences between two groups of children with spastic diplegia who received different intensities of early onset physiotherapy followed for 5 years. Brain Dev 2004;26(2):118-26.
Senthilkumar S, Swarnakumari P. A study on Vojta therapy approach to improve the motor development of cerebral palsy children. Int J Med Sci 2011;4(1&2):39-45.
Laufens G, Poltz W, Buchstein G, Schmiegelt F, Stempski S. [Improvement in locomotion by combined treadmill/Vojta physiotherapy applied to selected MS patients] [Article in German]. Phys Rehab Kur Med. 1999; 9: 187-189.
Hok P, Opavský J, Kutín M, Tüdös Z, Kaňovský P, Hluštík P. Modulation of the sensorimotor system by sustained manual pressure stimulation. Neuroscience. 2017.21;348:11-22.
Sanz-Esteban I, Calvo-Lobo C, Ríos-Lago M, Álvarez-Linera J, Muñoz-García D, Rodríguez-Sanz D. Mapping the human brain during a specific Vojta’s tactile input: the ipsilateral putamen’s role. Medicine (Baltimore). 2018;97(13):e0253. doi:10.1097/MD.0000000000010253
Hok P, Opavský J, Labounek R, et al. Efectos diferenciales de la estimulación por presión manual sostenida según el sitio de acción. Frente Neurosci . 2019; 13: 722. Publicado el 17 de julio de 2019 doi: 10.3389 / fnins.2019.00722
Zehr EP, Duysens J. Regulation of arm and leg movement during human locomotion. Neuroscientist. 2004 Aug;10(4):347-61.Ayala F, Cela CJ. Evolución Humana. Alianza Editorial 2013.
Dietz V, Fouad K, Bastiaanse CM. Neuronal coordination of arm and leg movements during human locomotion. Eur J Neurosci. 2001 Dec;14(11):1906-14.
Perales-López, L. El pisiforme, una nueva zona de desencadenamiento segmental de los contenidos cinesiológicos de la mano en la terapia Vojta. Aplicaciones en rehabilitación. Fisioterapia, 2013. Vol 35, (5), P 189-196.
Vojta V, Schweizer E. El descubrimiento de la motricidad ideal. Ediciones Morata 2011.
Sharma A. Transgenerational epigenetics: Integrating soma to germline communication with gametic inheritance. Mech Ageing Dev. 2017;163:15-22. doi:10.1016/j.mad.2016.12.015
Legoff L, D’Cruz SC, Tevosian S, Primig M, Smagulova F. Transgenerational Inheritance of Environmentally Induced Epigenetic Alterations during Mammalian Development. Cells. 2019;8(12):1559. Published 2019 Dec 3. doi:10.3390/cells8121559
Nicoglou A, Merlin F. Epigenetics: A way to bridge the gap between biological fields. Stud Hist Philos Biol Biomed Sci. 2017;66:73-82. doi:10.1016/j.shpsc.2017.10.002
Rao R, Jackson S, Howard R. Reflejos primitivos en la enfermedad cerebrovascular: un estudio comunitario de personas mayores con accidente cerebrovascular y estenosis carotídea. Psiquiatría Int J Geriatr . 1999; 14 (11): 964-972
Kapandji, A. I. (2006). Fisiología articular, Tomo 3, Editorial Médica Panamericana.
Perales L, Pérez AM, Atim MA, Varela E. Efecto de la terapia Vojta en la rehabilitación de la marcha en dos pacientes adultos con dano˜ cerebral adquirido en fase tardía. Fisioterapia. 2009;31:151—62.
Husárová, R. The vojta approach in adult patients. Rehabilitacia. 2005;42(3).
Cerling TE, Levin NE, Quade J, et al. Comment on the paleoenvironment of Ardipithecus ramidus. Science. 2010;328(5982):1105. doi:10.1126/science.1185274
Bramble DM, Lieberman DE. La carrera de resistencia y la evolución del Homo. Naturaleza . 2004; 432 (7015): 345-352. doi: 10.1038 / nature03052
Ambrosio Garcia Leal. El azar creador. Ed. Tusquets Editores. 2013
Judith Rich Harris. No hay dos Igaules. Funnanbulista 2015
Thompson NE, Demes B, O’Neill MC, Holowka NB, Larson SG. Surprising trunk rotational capabilities in chimpanzees and implications for bipedal walking proficiency in early hominins. Nat Commun. 2015 Oct 6;6:8416
Sizer, P.S., Jr., J.M. Brismee, and C. Cook, Coupling behavior of the thoracic spine: a systematic review of the literature. J Manipulative Physiol Ther, 2007. 30(5): p. 390-9.
Lee, L.J., M.W. Coppieters, and P.W. Hodges, Differential activation of the thoracic multifidus and longissimus thoracis during trunk rotation. Spine (Phila Pa 1976), 2005. 30(8): p. 870-6.
Edmondston, S.J., et al., Influence of posture on the range of axial rotation and coupled lateral flexion of the thoracic spine. J Manipulative Physiol Ther, 2007. 30(3): p. 193-9.
Kiernan D, O’Sullivan R, Malone A, O’Brien T, Simms CK. Pathological Movements of the Pelvis and Trunk During Gait in Children With Cerebral Palsy: A Cross-Sectional Study With 3-Dimensional Kinematics and Lower Lumbar Spinal Loading. Phys Ther. 2018;98(2):86-94. doi:10.1093/ptj/pzx113
Swinnen E, Goten LV, De Koster B, Degelaen M. Thorax and pelvis kinematics during walking, a comparison between children with and without cerebral palsy: A systematic review. NeuroRehabilitation. 2016;38(2):129-146. doi:10.3233/NRE-161303
G. Laufens, S. Seitz, G. Staenicke, Fachbereich. Fundamentos biológicos comparativos respecto a la locomoción innata, en particular respecto a la “reptación refleja” según Vojta. Fisioterapia 43. Jg (5/1991), Pag. 448-156.9.
Anexo 1:La paradoja de Thompson
¿Como sabemos que la inserción torácica de las cadenas musculares espirales son inestables en otros primates durante la locomoción bípeda?
Un estudio de Thompson (2015) analizó con tecnología de captación de imágenes la marcha bípeda en chimpancés y humanos revelando una aparente paradoja. Resulta que humanos y chimpancés caminamos con el mismo patrón cruzado (avanzamos pierna y brazo contrario a la vez) pero lo hacemos con un movimiento rotacional entre cintura pélvica y escapular diferente. Mientras que los humanos hacemos giros opuestos (desfase), los chimpancés giran en el mismo sentido (en fase).
Tomado de Thompson 2015
Entonces ¿como es posible que tengamos el mismo patrón cruzado? con ambas cinturas girando en el mismo sentido sería esperable que al caminar los chimpancés y otros simios avanzasen las extremidades del mismo lado al tiempo… pero no es así.
El movimiento rotacional de la sección torácica de la columna vertebral (CV) analizado en este estudio se enmarca en el plano transversal, pero la respuesta a esta contradicción se encuentra en otro plano: el frontal, (no incluido en el estudio) donde se producen los movimientos de inclinación lateral. Un plano con una influencia directa en los movimientos del plano transversal de la CV (Sizer 2007), sobre todo cuando la Musculatura autóctona de la CV (Multifidus y longisimus) no es capaz de sostenerla en su eje longitudinal (Lee 2005)
Entonces, la inclinación lateral del tronco conlleva una rotación compensatoria del lado contrario, principalmente desde una posición extendida de la CV (Edmondston 2007) como es el caso de la CV durante la marcha erguida. Esta es la rotación que Thompson ha registrado en su estudio, un subproducto o secuela de la claudicación que no guarda relación con la orientación dorsal de los iliacos, sino con la imposibilidad de su musculatura autóctona para sostener la CV en su eje longitudinal en la fase de apoyo monopodal de la marcha.
En resumen, si observamos la fase de apoyo monopodal en la marcha del chimpancé cuando apoya el pie izquierdo, su tronco se inclina hacia la izquierda, por tanto, su CV torácica rotará hacia el lado contrario (la derecha), el mismo lado hacia el que rota la hemipelvis izquierda que en ese momento esta en el apoyo (imagen de abajo derecha). Mientras, para el mismo apoyo izquierdo en el hombre, al mantenerse la CV enderezada en su eje axial, no existe inclinación lateral y por tanto el movimiento rotacional se expresa sin los condicionamientos biomecánicos que produce el movimiento en el plano frontal (imagen de abajo izquierda).
Modelo en desfase humano y modelo en fase del chimpancé.Tomado de Thompson 2015
De este modo se explica la supuesta incoherencia. Lo importante aquí es, que a pesar del handicap que supone la incapacidad de la musculatura autóctona para sostener la CV enderezada en el momento más complejo de la marcha (apoyo monopodal), el patrón cruzado se expresa igualmente en el chimpancé. Esto revela que el esquema rotacional entre cinturas en la misma fase que muestra el chimpancé en el estudio de Thompson no es una forma alternativa de ejecutar esta acción por sus particularidades anatómicas, sino la expresión de una compensación disfuncional para la marcha bípeda.
Evolución de movimiento de la CV en el plano frontal. Tomado de Laufens 1991
El modelo rotacional en desfase del humano es el marco de referencia para la marcha bípeda porque consigue mantener la CV en su eje axial del mismo modo que lo hacen los mamíferos terrestres cuadrúpedos en su segmento torácico (los reptiles mueven su CV en el plano frontal). En consecuencia, se trata del modelo eficiente seleccionado que el chimpancé puede ejecutar en su locomoción cuadrúpeda, pero no en la bípeda. Esto no depende de la orientación de los iliacos, sino de la capacidad de la musculatura autóctona para sujetar dinamicamente la CV.
Los estudios cinesiológicos en niños con lesiones neurológicas que comprometen la función de la musculatura autóctona, como es el caso de la parálisis cerebral (que no afecta a la inervación ni a la estructura sino a la función) muestran movimientos del tronco durante la marcha (Swinnen 2016, Kiernan 2018) equiparable a los que realizan los chimpancés, a pesar de hacerlo con unos iliacos plenamente humanos.
Típica inclinación lateral del tronco en la marcha en la parálisis cerebral
Durante la ontogenesis postural, en los primeros pasos que da el niño hacia delante, la musculatura autóctona se muestra temporalmente incapaz de controlar y sostener su columna en el eje axial. Entonces se puede observar las inclinaciones laterales del tronco (Vojta 2011) que recuerdan el mismo movimiento de otros primates al caminar.
Lactante de 13 meses sano dando sus primeros pasos con una marcada utilización del plano frontal
Anexo 2: El patrón de la 1ª posición
Posición de partida del patrón de la 1ª posición. El ejercicio debe hacerse al menos una vez con la cabeza girada de cada lado, por lo que es indiferente el lado de comienzo
El patrón de la 1ª posición descrito por Vojta (Vojta 1972) se inicia desde una posición similar a la postura fetal con la cabeza girada hacia un lado y con el brazo de ese mismo lado flexionado hacia delante (ver imagen) y finaliza con la puesta en bipedestación e inicio de la marcha. Todo ello de forma refleja presionando cualquiera de las zonas descritas por Vojta (ver imagen) de forma individual o en combinación. El patrón completo toma el nombre de la postura con la que se inicia.
Desde esta 1ª posición hasta la bipedestación, el patrón pasa por diferentes posiciones, con contenidos cinesiológicos específicos. Para obtener un mejor aprovechamiento terapéutico de estos contenidos Vojta decidió fragmentar este patrón en séis partes, cada una de las cuales tiene su posición de partida específica (imagen de abajo).
Por tanto, no es necesario para alcanzar el final del patrón (bipedestación y marcha) comenzar la activación desde la 1ª posición, se puede hacer desde cualquiera de las seis posiciones. Es más, ya desde la 1ª posición es posible acceder a muchos componentes cinesiologicos propios de la bipedestación y marcha como es la extensión axial de la CV, la retroversión de la pelvis y la rotación externa de la articulaciones esféricas.
Considere las diferentes posiciones de este patrón como las etapas del camino de Santiago. Todas pertenecen al mismo camino continuo pero cada etapa ofrece paisajes y ambientes que aportan experiencias distintas. Si decidimos hacer una etapa concreta es porque valoramos lo que aporta ese tramo a nuestros intereses dentro del conjunto del camino. Empezar el camino muy cerca del objetivo solo te servirá si el resto ya lo has andado, en caso contrario no te servirá. Del mismo modo poner a caminar a un niño que no es capaz de controlar su cabeza y tronco en posturas más básicas tampoco será de ayuda. Un bebé sano al nacer no tiene como objetivo caminar, sino que caminar es la consecuencia de la maduración de su SNC a lo largo de un periodo próximo al año.
La terapia Vojta utiliza este patrón como instrumento para la rehabilitación de la marcha (en combinación con otros patrones de la Locomoción refleja), aunque no la aborda directamente, si no a partir de la activación de los contenidos cinesiologicos parciales que la integran desde sus componentes básicos.
Si en esta metáfora el camino es un constructo que forma parte de nuestra historia cultural, el patrón innato de la primera posición, descubierto por Vojta, forma parte de nuestra historia evolutiva.
Fotogramas de una secuencia real de activación refleja del patrón de la 1ª posición en un adulto sano desde la postura inicial (postura de partida de la 1ª posición) hasta la final
La adquisición de la marcha bípeda independiente constituye uno de hitos más importantes en el proceso de habilitación/rehabilitación en niños y adultos. Cualquier método de tratamiento fisioterápico tiene el reto de demostrar su validez y competencia en este hito esencial de nuestro desarrollo motor. Para conocer las aportaciones de la terapia Vojta a la rehabilitación de la marcha antes es necesario revisar cómo llega el bebe a conseguir caminar de forma libre durante la ontogénesis, qué patrones son claves y de donde proceden. A continuación analizaremos las características de los patrones innatos descritos por Vojta, para finalmente establecer cuales son las relaciones entre ambos. Solo así podremos establecer de forma rigurosa cómo la terapia Vojta contribuye a la habilitación o rehabilitación de la marcha bípeda.
La marcha humana
Se trata del patrón de locomoción más estudiado, pero ha sido en las últimas décadas, gracias a la aparición de las nuevas tecnologías, cuando hemos podido describir en detalle esta insólita manera de desplazarnos como mamíferos terrestres sobre nuestras patas traseras principalmente en la marcha del adulto (1-3). Se trata de análisis de la cinemática y la cinética de la marcha que no consideran la marcha como el ensamblaje de diferentes patrones posturales en un proceso programado genéticamente en constante interacción con el medio y subordinado a la maduración cognitiva (4). En definitiva, no se contempla la procedencia de los patrones que la integran, sino el resultado final (Figura 1).
Figura 1: Representación del los principales Items de la ontogenesis que aportan patrones básicos al patrón de marcha
1.1 Enfoques terapéuticos en la habilitación/rehabilitación de la marcha
¿Y porque habrían de hacerlo? si la marcha es el contenedor integrado de todos los patrones que le han precedido, se entiende que, abordando directamente el entrenamiento del patrón final, quedarían asimilados todos sus patrones subyacentes. Actualmente éste es el enfoque que acumula más evidencia (5,6), y que se puede resumir en que el entrenamiento continuado de una función mejora dicha función(7). Si bien, se trata de mejoras en variables cuantitativas como la velocidad en cortas distancias(8), y la resistencia, y no se informa de la calidad en la ejecución ni del sostenimiento en el tiempo de los resultados(9). Corr apunta que detrás de éstos resultados positivos está la mejora en la fuerza de las piernas (10,11), no de la coordinación de los patrones implicados, o de la calidad de su cinesiología… de la fuerza, un factor compensatorio necesario cuando se entrena una función que exige una actividad antigravitatoria para la cual se carecen de los patrones innatos genéticos (y por ende eficientes) implicados en la marcha (12).
Niño con diparesia caminando
Veamos un ejemplo: En un niño con diparesia espástica que camina de forma independiente la retroversión de la pelvis está ausente. En su defecto la pelvis se sostiene en una primitiva anteversión, una situación que condiciona toda la biomecánica global, empezando por la limitación de la rotación externa y la extensión de la cadera(13). Un hecho que afectará en cascada a la biomecánica de todo el miembro inferior, donde los aductores tomarán un papel disfuncional al no actuar de forma coordinada con sus sinergistas, rotando internamente las caderas y aproximándolas de forma que ambas rodillas lleguen incluso a contactar durante la marcha. Fracasando, además, en su función de antigravitación en el plano frontal. El entrenamiento de la marcha en estos niños, lejos de activar el componente ausente (la retroversión) se «perfecciona» en el uso de los mecanismos compensatorios incrementando progresivamente sus componentes parciales: flexión del tronco, cadera, rodilla y rotación interna de cadera, entre otros(14). En esta situación la retroversión no aparece con el entrenamiento de la marcha, sino que se aleja aún más de ella.
2. El patrón cruzado en la locomoción.
En las monografías que tratan la marcha (1-3) apenas se profundiza en el movimiento de braceo del miembro superior. Un enfoque que ha influido en el diseño de dispositivos robóticos y exoesqueletos que no consideran ni estimulan el movimiento de balanceo de los brazos, tampoco en los robots pediátricos(15). Una situación que se contradice con las bases neurofisiológicas de la locomoción cruzada que nos caracteriza como vertebrados (16).
Así, la explicación del movimiento de contra rotación entre las cinturas escapular y pélvica tiene diferentes hipótesis, mientras unos lo justifican como un mecanismo de compensación para absorber las fuerzas generadas a causa de nuestra marcha bípeda, otros sostienen que los brazos forman parte indisoluble del patrón de locomoción aportando una pequeña parte de energía al proceso de marcha(17). Por tanto, de la revisión de la evidencia científica en esta materia se podría deducir que la marcha bípeda es competencia casi exclusiva de las piernas, lo cual se contradice con un estudio de Huang(18) que demuestra la fuerte influencia del Core en la marcha de niños con PC. No obstante, en el estudio de la marcha normal se mantiene el papel secundario del tronco, y la columna vertebral apenas se menciona más allá de su función de mantener erguido el tronco mientras se mueven las piernas. Prueba de ello es el nombre con el que se refieren en la literatura al tren superior: «miembro pasajero»(19) ya que se considera que es una estructura transportada.
2.1. La columna vertebral en la locomoción con el patrón cruzado.
La columna vertebral como sistema preestablecido, autoactivo y altamente dinámico, es la clave de control de cualquier patrón innato de locomoción de cualquier vertebrado(20). Con la locomoción en posición vertical, y sus movimientos cíclicos en los tres planos del espacio, la construcción de la columna vertebral humana no es comparable con la de ninguna de los cuadrúpedos(21). La diferenciación funcional de la compleja musculatura autóctona está en estrecha interconexión con la diferenciación funcional de toda la musculatura del cuerpo, y es responsable del control intersegmentario de la columna vertebral (CV), y por consiguiente, también de los movimientos rotacionales vertebrales que subyacen al giro entre cinturas durante la marcha(22).
Este movimiento de la CV forma parte indisoluble del patrón de locomoción cruzado en coordinación con las extremidades(23). Por tanto, su activación debería tomarse en cuenta en cualquier proceso terapéutico de la marcha. Teniendo esto presente, para entender mejor la naturaleza del patrón cruzado antes es necesario analizar cómo se comporta la CV durante la ontogénesis en los diferentes patrones de locomoción.
2.1 a) El arratre
Es el primer patrón de locomoción y surge a mediados del tercer trimestre. Una locomoción dependiente de los brazos que utiliza como base un patrón ya conocido por el bebe: el patrón asimétrico en un codo. En el arrastre la CV se comporta contra rotando a nivel de la charnela dorso-lumbar, lo cual pertenece al patrón homólogo de locomoción(24). Su duración es corta, no constante y variable en sus formas de expresión.
2.1. b) El gateo
El siguiente patrón de locomoción es el gateo, (finales del tercer trimestre) donde se dan dos formatos en función de la velocidad: (a) con tres puntos de apoyo, más lento, y donde se produce una contra rotación en la charnela dorso lumbar (Figura 2), y (b) con dos puntos de apoyo, más rápido, y donde toda la CV rota en la misma dirección (Figura 3). El gateo, es pues, una locomoción mixta que combina el formato cruzado y homologo (24).
Figura 2: Secuencia del gateo a 3 puntos donde se alternan patron cruzado y homologo. Observe que al patron cruzado se le reconoce por la forma geometrica del tronco en trapecio, mientras que el patron homologo describe una forma en paralelogramo.
Figura 3: Secuencia del gateo en 2 puntos, propio del patron cruzado identificable por la figura geometrica del trapecio alterno.
Aunque poco frecuente, en el gateo también puede observarse el patrón homólogo como forma hegemónica de locomoción donde las extremidades del mismo lado se mueven a la vez produciéndose una contra rotación en la charnela dorso-lumbar, igual que en el gateo a tres puntos. Se trata de un comportamiento limitado en el tiempo, ya que no se prolonga más allá del 5º trimestre cuando el bebé se lanza a la marcha bípeda. Entonces, el patrón homologo queda subordinado al patrón cruzado, lo que significa que, aunque dejamos de verlo, su acción se sigue ejerciendo sobre la CV en momentos concretos de la marcha como son los cambios de dirección, cuando nos detenemos, o caminamos muy lento.
Anexo 1: ¿Qué pasa con los niños sanos que no gatean?
Se estima que en torno al 18% de los bebes con un desarrollo sano no gatean, empleando diferentes variantes para sus desplazamientos antes de ponerse a caminar(25). ¿Significa esto que no disponen del patrón cruzado hasta que comienzan a andar? ¿es el patrón cruzado producto del aprendizaje? Las razones por las cuales un bebe se salta la etapa del gateo no están claras, pero eso no implica que carezcan del patrón cruzado antes de caminar ya que está presente en otros patrones básicos que sí deben estar, como la sedestación oblicua que deriva en la sedestación bisquiatica (manos libres), y de forma generalizada en la marcha lateral que los niños realizan apoyándose con las manos en los muebles.
Curiosamente, aquellos que no han gateado de forma ortodoxa antes de caminar lo hacen, sin excepción, posteriormente sin entrenamiento previo. Por lo que puede afirmarse que el patrón cruzado de la marcha bípeda «depura» y corrige cualquier patrón alternativo o sustitutivo que el bebé hubiese fijado como forma de locomoción previo a la marcha. Por esta razón no debe confundirse el gateo con el patrón cruzado, que también puede realizarse con un patrón homologo. Por tanto, carece de sentido el entrenamiento del gateo como requisito necesario para la marcha, ya que los niños sanos que no gatean disponen del patrón cruzado, aunque no lo utilicen para gatear y no presentan más dificultades que los gateadores para caminar en tiempo y forma(26).
Así, con el objetivo en la habilitación de la marcha en AT tiene más sentido centrarse en la identificación de los déficits cinesiológicos del patrón cruzado que comienzan a expresarse a mediados del 2º trimestre con el patrón del apoyo asimétrico en un codo, y que continúan diferenciándose con el volteo (finales del 2 trimestre) y la sedestación oblicua (mitad del tercero), hasta su expresión locomotora con el gateo y la marcha lateral como antecedentes de la marcha libre. A finales del tercer trimestre y principios del cuarto, la gran variedad de patrones posturales y motrices disponibles dejarán al lactante margen de maniobra suficiente para imprimir su carácter en la motricidad. A ello contribuyen factores internos como la personalidad, que le llevan a tomar elecciones según sus preferencias, condicionadas, o no, por disfunciones posturales transitorias que puedan orientar una postura asimétrica «fijada» por el uso, hasta su corrección por la imposición del patrón cruzado en la marcha, y que compartimos con el 95% de los vertebrados terrestres(27).
También los factores ambientales (entorno afectivo y social) jugaran un papel importante en sus elecciones motrices. En consecuencia, es el análisis de la maduración cinesiológica del patrón cruzado durante la ontogénesis y no el gateo (que como forma de locomoción transitoria tan solo es una expresión más del patrón cruzado en la ontogénesis postural) lo que resulta sustancial en la valoración de los déficits motores del niño. De hecho, los niños con diparesia llegan a realizar distintas formas de gateo, lo que no implica que lo ejecuten con la cinesiología propia del patrón cruzado. Con esto no se pretende restar importancia al gateo, por cuanto evidencia la viabilidad y disponibilidad del patrón cruzado para la locomoción bípeda, tan solo ponerlo en el amplio y complejo contexto del desarrollo motor. Como decía el neurólogo Henner, mentor de Vojta: «Haz caso de la cinesiología y solo a través de ella conocerás al SNC»)
2.1. c) La marcha bípeda
En la locomoción bípeda se produce un movimiento de contra rotación entre las cinturas escapular y pélvica, lo cual puede hacer pensar que la CV se comporta del mismo modo al caminar. Sin embargo, esta idea no considera que en la articulación sacroilíaca durante la fase de apoyo monopodal se produce un movimiento de contra nutación haciendo que la cara ventral del sacro se oriente hacia el iliaco de la pierna de apoyo(28), es decir, la columna lumbar tiende a rotar en la misma dirección que la columna dorsal.
Figura 4: Rotación de la CV en la misma dirección en la fase de apoyo monopodal
En otras palabras, durante la fase de apoyo monopodal la CV rota en la misma dirección, justo cuando se expresa con mayor nitidez el patrón cruzado en la marcha (Figura 4). Una situación análoga a la que vimos en la locomoción cuadrúpeda cruzada en dos puntos de apoyo del gateo. En cambio, en la fase de apoyo bipodal, la columna lumbar se posiciona en la línea media con la columna dorsal girando hacia el brazo que avanza. Se trata de un instante donde se produce un movimiento de contra rotación relativo, un esquema que pertenece al patrón de locomoción homólogo que también se produce en el gateo a 3 puntos.
Luego gateo y marcha bípeda responden a programas biomecánicos análogos en relación al movimiento de la CV. ¿Significa ésto que el gateo es una condición necesaria para desarrollar la marcha bípeda? No, pero si lo es disponer tanto de la cinesiología del patrón cruzado como del homólogo en la CV en coordinación con las extremidades(29).
2.2. El patron cruzado en la terapia Vojta
Dentro de la LR de Vojta es el patrón de la Reptación refleja (RR) (Figura 5), el que mejor representa el patrón cruzado, ya que los apoyos se encuentran predefinidos desde la posición de partida en una diagonal, mientras que la otra realiza el movimiento de avance(30). El análisis de la cinesiología del patrón de la RR revela que la CV gira en la misma dirección, lo mismo que sucede en el gateo cruzado a dos puntos y en la fase monopodal de la marcha(31). Distintos formatos de locomoción con un denominador común: el patrón cruzado, controlado por los mismos circuitos neurales como ha demostrado el neurofisiólogo Volker Diezt(32). Probablemente
Figura 5: Posición de partida del patron cruzado de la reptción refleja
Esta estrecha relación de los patrones neurales implicados explicaría la transferencia de la activación del patrón de la RR al patrón de marcha en niños y adultos, sin necesidad de entrenarla de forma específica y además, activada en posiciones en decúbito (33-36). Una vinculación (marcha y RR) que fue investigada por Laufens en pacientes con Esclerosis múltiple, resultando más efectiva para la marcha caminar 7 minutos antes de realizar la TV que su abordaje independiente(37). Combinación confirmada por Reumont en pacientes con lesión medular incompleta (38).
Empleando RMN funcional Hok(39) demostró cómo desde el estímulo en la zona del talón se producía neuromodulación en la Formación reticular pontomedular correspondiente a una función solicitada en la mano contralateral. Una estructura competente en la activación de los GCPs (Generadores Centrales de Patrones). Un hecho que ha sido contrastado utilizando EMG, demostrando la existencia de una secuencia rítmica temporo-espacial compatible con la activación del patrón cruzado en la RR(40,41), análoga, y por tanto transferible a la que se expresa durante la marcha(23).
En consecuencia, con la aplicación terapéutica de las locomociones reflejas innatas es posible estimular de forma refleja y directa a la musculatura autóctona axial, una estructura nuclear en la ejecución de los movimientos rítmicos generados por el GCPs., en relación a las extremidades,y expresados en coordinaciones innatas en la CV (patrón cruzado y homologo), que están presentes en diferentes fases de la locomoción bípeda(42-48)
3. Contenidos cinesiologicos analogos entre la LR y la ontogenesis relacionados con la marcha.
El estudio y análisis de la cinesiología del desarrollo fue consecuencia y no causa del descubrimiento de las locomociones reflejas(24). Así es como Vojta y colaboradores comprobaron que los patrones globales que había descubierto y descrito a lo largo de 18 años de investigaciones contenían los patrones parciales implícitos en el patrón cruzado de la marcha bípeda, incluidos los tres requisitos de toda locomoción: el control postural automático, reacciones de enderezamiento sobre puntos de apoyo y movimiento fásico segmental propositivo(26).
Por tanto, la realización de los ejercicios de la TV con el objetivo de alcanzar, o mejorar la calidad del patrón de marcha, no pretenden la adquisición del volteo o la reptación (como se confunde habitualmente por la denominación de los patrones reflejos descubiertos), si no la activación refleja de los patrones parciales que pertenecen al patrón cruzado de la marcha, y que están ausentes o incompletos en los niños con alteraciones del desarrollo motor.
Estos patrones parciales de la LR, análogos a los patrones parciales que observamos en la ontogénesis (Figura 6), y que más directamente guardan relación con el patrón cruzado de la marcha(28) son:
Extensión axial de la CV,
Retroversión pélvica,
Rotación externa de las articulaciones esféricas,
Diferenciación de la función muscular en el tronco (autóctona de la CV y Core), y en las extremidades, donde se configuran puntos de apoyo en las partes distales.
Patrón cruzado y homólogo en la CV.
Enderezamiento del tronco sobre puntos de apoyo.
Desplazamiento del centro de gravedad.
Figura 6:Representación de los patrones globales de la ontogénesis que contienen los patrones parciales presentes en la LR
4. Conclusiones
Los patrones de la LR de Vojta contiene todos los patrones parciales esenciales que pertenecen a la marcha bípeda, patrones preprogramados en el genoma y que tienen su origen en nuestra herencia evolutiva. Su accesibilidad para imprimir en el cerebro del bebé patrones de la ontogénesis ideal que conducen hacia el enderezamiento y la marcha, junto con la facilidad para su reproducción por parte de la familia hacen de la TV una herramienta de uso imprescindible no solo en Atención temprana, también en adultos con daño neurológico adquirido, u otras patologías que comprometen a la marcha.
(Anexo 2: La TV un instrumento de diagnóstico temprano.
La validación de los patrones de la ontogénesis como referencia comparativa para determinar el grado de madurez motora del niño en relación a su edad cronológica ha permitido disponer de un instrumento eficaz para evaluar el progreso con una terapia determinada(49). Cada uno de estos patrones o hito del desarrollo fue deconstruido en sus componentes parciales de modo que Vojta y colaboradores pudieron reconocer e identificar las novedades cinesiológicas que debutaban en cada etapa del desarrollo(23). Resultó que los tiempos de aparición eran muy similares en la mayoría de los niños sanos independientemente de su procedencia. Esta pauta regular apuntaba a una programación genética. La confirmación de esta programación de los patrones posturales llegó con la implementación de las reacciones posturales. Una serie de 7 pruebas diagnósticas que demostraban la regularidad de las respuestas automáticas globales ante la aplicación de determinados estímulos(50). Las mismas respuestas se repetían en edades cronológicas similares, respuestas que se iban modificando siguiendo tiempos regulares desde el nacimiento hasta alcanzar la marcha, y cuyos patrones parciales se correlacionaban temporalmente con los que aparecían en la motricidad espontanea(51). Por esta razón su batería de Test posturales puede determinar la presencia de alteraciones en la coordinación central en cualquier periodo del desarrollo desde una edad muy temprana(52). En caso de observar signos anormales en estos Test, la aplicación temprana de terapia es lo indicado en prevención de que patrones sustitutivos anormales queden fijados por el uso.)
(Anexo 3: Patrones de la evolución.
La programación genética de las respuestas posturales y motrices de las reacciones posturales así como la existencia de tiempos de maduración regulares no solo estaban presentes en nuestra especie, resulta que son compartidas por todos los primates como demostró la primatóloga Hideko Takeshita, aplicando los mismos test a 6 especies de primates diferentes, incluido la nuestra(53). Las diferencias encontradas se relacionan con los matices anatómicos específicos de cada especie (chimpancés y gorilas carecen de flexión dorsal de muñeca) pero sobre todo con los tiempos de maduración en alcanzar cada hito, que se acortan a medida que la especie de primate se aleja más de nuestra línea filogenética. En consecuencia, son los chimpancés los que mayores similitudes presentan en tiempo y forma a las reacciones posturales descritas por Vojta. En otras palabras, los humanos somos los que nacemos más inmaduros entre los primates, lo cual nos brinda un tiempo muy valioso para el aprendizaje y la maduración necesaria para alcanzar el complejo patrón de locomoción bípeda(54).
Bibliografia
1.Plas F, Viel E, Blanc Y. “La marcha humana”. Barcelona: Masson; 1996
2. Cámara J. Análisis de la marcha: sus fases y variables espacio-temporales. 2011;7(1): 160-173.
3. Perry J, Burnfield JM. Análisis de la Marcha. Ed Base. 2015.
4. Zelazo PR. The development of walking: new findings and old assumptions. J Mot Behav 1983; 15:99-137
7. Anne Shumway-Cook, Marjorie H. Woollacott. (1995). Control Motor. Williams & Wilkins.
8. Noelle G Moreau, Amy Winter Bodkin, Kristie Bjornson, Amy Hobbs, Mallary Soileau, Kay Lahasky. Effectiveness of Rehabilitation Interventions to Improve Gait Speed in Children With Cerebral Palsy: Systematic Review and Meta-analysis. Phys Ther. 2016 Dec;96(12):1938-1954. doi: 10.2522/ptj.20150401.
9. Cimolin V1, Galli M, Piccinini L, Berti M, Crivellini M, Turconi AC. Quantitative analysis of gait pattern and energy consumption in children with cerebral palsy. Appl Biomater Biomech. 2007 Jan-Apr;5(1):28-33.
10, Corr BB, Stuberg WA, Arpin DJ, Kurz MJ. Changes in lower extremity strength may be related to the walking speed improvements in children with cerebral palsy after gait training. Res Dev Disabil. 2018 Feb;73:14-20.
15. Swinnen E, Beckwée D, Meeusen R, Baeyens JP, Kerckhofs E. Does robot-assisted gait rehabilitation improve balance in stroke patients? A systematic review. Top Stroke Rehabil. 2014 Mar-Apr;21(2):87-100. doi: 10.1310/tsr2102-87.
16. Grillner S, Wallén P: Central pattern generators for locomotion, with special reference to vertebrates. Annu Rev Neurosci, 1985, 8: 233–261.
17. Collins SH, Adamczyk PG, Kuo AD. Dynamic arm swinging in human walking. Proc Biol Sci. 2009 Oct 22;276(1673):3679-88.
19. Molina, Carratalá. La marcha humana. Biomecanica, evaluación y patologia. Ed Panamericana. 2020.
20. Griller S, Buchanan J, Kallen P, Brodin L. 1988. Neural Control of locomotion in lower vertebrates. In: Cohen AH, Rossignol S, Grillner S, (Eds) Neural control of rhythmic movements in vertebrates, 1-40. New York.
21. Lovejoy C. Owen Historia natural de la marcha y de la postura humanas: columna vertebral y pelvis. EMC (Elsevier SAS, Paris), Kinesiterapia – Medicina física, 26-007-B-10, 2005.
22. Zehr EP, Duysens J. Regulation of arm and leg movement during human locomotion. Neuroscientist. 2004 Aug;10(4):347-61.Ayala F, Cela CJ. Evolución Humana. Alianza Editorial 2013.
23. Dietz V, Fouad K, Bastiaanse CM. Neuronal coordination of arm and leg movements during human locomotion. Eur J Neurosci. 2001 Dec;14(11):1906-14.
24. Vojta V, Schweizer E. El descubrimiento de la motricidad ideal. Ediciones Morata 2011.
25. Fernández-Jaén A, Martín Fernández-Mayoralas D, Calleja-Pérez B, Muñoz-Jareño N. Detección y manejo del retraso psicomotor en la infancia. Pediatr Integral. 2011:15:753
26- Martín Fernández-Mayoralas D, Fernández-Jaén A, Muñoz-Jareño N, Calleja-Pérez B. Valoración del desarrollo psicomotor normal. Pediatr Integral. 2011;15:7
27. Adria Casinos. La Locomoción. CSIC. 2018.
28. Ricard F. Tratamiento osteopático de las algias lumbopélvicas. 2005. Ed Médica Panamericana.
29. Dietz V, Fouad K, Bastiaanse CM. Neuronal coordination of arm and leg movements during human locomotion. Eur J Neurosci. 2001 Dec;14(11):1906-14.
31. Vojta V. Peters A. El principio Vojta. Springer-Verlag Ibérica 1995
32. Dietz V. Quadrupedal coordination of bipedal gait: implications for movement disorders. J Neurol. 2011 Aug;258(8):1406-12.
33. Husárová, R. The vojta approach in adult patients. Rehabilitacia. 2005;42(3).
34. Perales L, Pérez AM, Atim MA, Varela E. Efecto de la terapia Vojta en la rehabilitación de la marcha en dos pacientes adultos con dano˜ cerebral adquirido en fase tardía. Fisioterapia. 2009;31:151—62.
35. Loaiciga Espeleta C. La terapia Vojta como herramienta de tratamiento de las alteraciones neuromotoras en la marcha de niños y adultos. Tesis Doctoral. Salamanca, 2014
36. Min Soo Sonb, Do Hee Junga, Joshua (Sung) H. Youa,∗, Chung Hwi Yic, Hye Seon Jeonc. Effects of dynamic neuromuscular stabilization on diaphragm movement, postural control, balance and gait performance in cerebral palsy. NeuroRehabilitation 41 (2017) 739–746.
37. Laufens, G, Jügelt, E, Poltz, W, Reimann G G. Ablauf und Erfolg einer Vojta Physiotherapie an ausgewählten MS-Patienten. Verhandlungen der Deutschen Gesellschaft für Neurologie. 1991;6:230-1.
38. Von Reumont AAbBPSiL. [Combination of treadmill and vojta physiotherapy after incomplete paraplegia] [German abstract] Abstract presented at the German Physiotherapeutic Congress September 2012 in Leipzig, Germany. 2012.
39. Hok P, Opavský J, Kutín M, Tüdös Z, Kaňovský P, Hluštík P. Modulation of the sensorimotor system by sustained manual pressure stimulation. Neuroscience. 21 de 2017;348:11-22.
41. Pavlu D, Véle F, HavlíˇckováL . Electromyography and kinesiological analysis of Vojta´s therapeutical principle. Rehab Fyzil. 2000;7:34-9
42. Kanda T, Yuge M, Yamori Y, Suzuki J, Fukase H. Early physiotherapy in the treatment of spastic diplegia. Dev Med Child Neurol 1984;26(4):438-44. 15.
43. Ha SY, Sung YH. Effects of Vojta method on trunk stability in healthy individuals. J Exerc Rehabil. 2016; 12: 542-547.
44. Kotnik SS. Vojta in rehabilitation of children with neurodevelopmental disorders. Paediatr Croat 2012;56(Supplement 1):227-31
45. Lim H, Kim T. Effects of vojta therapy on gait of children with spastic diplegia. J Phys Ther Sci. 2013; 25: 1605-1608.
46. Kanda T, Pidcock FS, Hayakawa K, Yamori Y, Shikata Y. Motor outcome differences between two groups of children with spastic diplegia who received different intensities of early onset physiotherapy followed for 5 years. Brain Dev 2004;26(2):118-26.
47. Senthilkumar S, Swarnakumari P. A study on Vojta therapy approach to improve the motor development of cerebral palsy children. Int J Med Sci 2011;4(1&2):39-45.
48. Laufens G, Poltz W, Buchstein G, Schmiegelt F, Stempski S. [Improvement in locomotion by combined treadmill/Vojta physiotherapy applied to selected MS patients] [Article in German]. Phys Rehab Kur Med. 1999; 9: 187-189.
49. Sanz Mengibar JM, Santonja-Medina F, Sanchez-de-Muniain P, Canteras-Jordana M. Can Clinical Assessment of Locomotive Body Function Explain Gross Motor Environmental Performance in Cerebral Palsy? J Child Neurol 2016; 31 (4): 474-480.
50. Costi, G, Radice, A, Raggi, A, Kron, A, Angrisano, E, Busato G et al. Vojta´s seven postural reactions for screening of neuromotorial diseases in infant, Research of 2308 cases. LA pediatrica Médica Chirurgica, (1983) Vol 5, nº 1-2.
51. Lajosi, F, Bauer H, Avalle C. Early diagnosis of central motor disturbances by postural reflexes after Vojta. XIV International Congress of Pediatrics, Barcelona 1980.
52. Vojta V. Early diagnosis and therapy of cerebral movement disorders in childhood. C. Reflexogenous locomotion – reflex creeping and reflex turning. C1. The kinesiologic content and connection with the tonic neck reflexes. Z Für Orthop Ihre Grenzgeb 1973;111(3):268-91.
53. Takeshita H, Tanaka, M, Matsuzawa T. Development of Postural Reactions in Infant Japanese Monkeys. The Annual of animal Psichology, 33 (1984)
54. Arsuaga JL, Martinez Mendizabal I. La especie elegida. Temas de Hoy.1998
En este Post se va a tratar un tema inédito en neurorehabilitación, el papel jugado por la musculatura autóctona de la columna vertebral en el control de la postura y el movimiento, tanto en la normalidad como en la patología, así como su enfoque, o ausencia de él, en las distintas metodologías que tratan estas patologías.
Estado de la cuestión
Tras un daño neurológico el 88% de los pacientes mantienen alteraciones funcionales persistentes en el miembro superior(MS) con un impacto negativo en su calidad de vida (Kwakkel 2013). El abordaje de la rehabilitación de estas secuelas, ha experimentado un importante auge con la implementación de las nuevas tecnologías como son los dispositivos robóticos (Amadeo, Diego, Pablo o Armeo), donde el MS parético es facilitado y guiado para la realización de movimientos orientados a ejecutar distintas acciones en contextos muy variados, como la manipulación de objetos reales, la ejecución de un programa de ejercicios pautados, o bien seguir un juego con fines terapéuticos (Dixit 2019).
No obstante, la evidencia científica de estos dispositivos no es concluyente, con trabajos que apoyan su eficacia (Sale 2014 y Aprile 2020) y otros que la cuestionan (Veebeek 2017 y Laffont 2019). Otro campo que ha tenido un gran desarrollo en la rehabilitación del MS es la realidad virtual, tanto de forma aislada como en combinación con dispositivos robóticos, que ha demostrado eficacia como actividad complementaria a la terapia convencional (Brunner 2017) aunque no se informa de efectos sobre el tronco y aún menos sobre la musculatura autóctona (MA) de la columna vertebral (CV).
Básicamente la rehabilitación del MS con estos dispositivos consiste en facilitar movimientos de alcance y manipulación del brazo parético en sedestación con el resto del cuerpo en actitud más o menos pasiva. La postura global, y más concretamente el tronco apenas es tenida en cuenta en los estudios que avalan la aplicación de estas tecnologías, o lo que es lo mismo, se ignora el papel de la MA de la CV, también conocida como paravertebral o epiaxial. Una musculatura que trabaja en sinergia con la musculatura ventral (abdominales) en el control postural del tronco que subyace a los movimientos del MS.
Si se considera que la atrofia de la musculatura del tronco (MA incluida) es más intensa que la producida en las extremidades afectadas en pacientes neurológicos (Chang 2019) sería interesante comprobar como estos procedimientos influyen en esta musculatura.
Precisamente una revisión sistemática de Alhwoaimel (2018) se preguntaba esto mismo ¿existe una correlación entre ejercitar el tronco con la mejoría de la función del MS parético? El resultado fue claro, los ejercicios para el tronco mejoraron el tronco pero de esta mejoría no se benefició el MS ¿Cómo podemos interpretar esto? que el tronco puede trabajar con independencia de las extremidades pero no a la inversa. En otras palabras, el MS trabaja en sincronía con el tronco, en concreto con la MA del segmento torácico de la CV.
Sin embargo, el trabajo a la inversa tiene mejores resultados como apunta un trabajo de Thrane (2018) donde la mejoría en las funciones del brazo autopercibidas se correlacionaron con los desplazamientos mejorados del tronco. De lo que se desprende que desde la extremidad se puede acceder a la activación automática de la MA.
Quizá los estudios recogidos en la revisión de Alhwoaimel sobre la no influencia del tronco en la mejoría de la función del MS han quitado el foco sobre el control postural que subyace a la movilidad del MS, mayoritariamente en sedestación. Tal vez el hecho de que cualquier adulto sano pueda manipular perfectamente desde una sedestación relajada (con cifosis), e incluso desde cualquier posición, puede llevar a pensar que esto es extensible a una persona que a perdido su capacidad para controlar su postura de forma automática después de un daño neurológico.
Sin embargo, en lesiones como la hemiparesia, donde se produce una especie de regresión en el control postural automático esto no es posible, ya que existe una desconexión en la postura axial con la extremidad afecta. Además, la musculatura ventral de este lado también se ve comprometida sumando más disfunción a la coordinación global de la postura en relación a las extremidades. Por contra, el lado sano, al no estar afectada su inervación provoca una actividad postural asimétrica, que refuerza el uso del lado sano, contribuyendo a una alteración de la representación somato-sensorial del lado afecto (Razmus 2017).
En la linea de potenciar la representación metal de una acción sensorio-motora como recurso en la rehabilitación postural y motriz, la imaginería motora ha realizado interesantes progresos mostrando mejoras significativas en diferentes estudios (Fernández 2018). Cabe suponer que estas mejoras en el uso del MS afecto llevan implícito una activación de la MA acorde con dichas mejoras, aunque no se informe específicamente de ello en estos trabajos.
Terapia restrictiva
Precisamente el éxito a corto plazo de otro enfoque, como es la terapia restrictiva (Yadav 2016) en las hemiparesias, incide en la activación sensorio-motriz del hemisferio dañado anulando la actividad del brazo sano. Esto provoca la activación del MS afecto incluido el hemicuerpo donde la MA esta integrada. Se trata en definitiva de una activación de la MA provocada por la movilidad fasica obligada del MS, eficaz por la elevada intensidad del programa de ejercicios y frecuencia de las sesiones. Factores que complican la adherencia al tratamiento, una posible causa de su débil evidencia a largo plazo como apunta una revisión sistemática con meta-análisis de Etoom (2017). Además, su aplicación en otras patologías neurológicas de afectación global (neurodegenerativas, TCE, o medulares) es más limitada cuando no imposible.
1.1 El circulo vicioso
En todas las lesiones neurológicas que afectan al control motor, la MA de la CV funciona de forma insuficiente o deja de hacerlo, provocando el fracaso del control postural en el órgano axial. Cuando su afectación se limita a un lado de la CV, como en la hemiparesia, (o formas asimétricas de la EM) la MA del lado sano es la responsable de provocar una inclinación hacia este lado, con el efecto asociado de rotación del cuerpo vertebral hacia el lado contrario: el afecto. Esto se traduce en que podemos observar una excurvación o convexidad en la zona alta del segmento dorsal de la CV.
Esta disfunción en el control del eje axial tendrá un efecto catastrófico sobre la diferenciación tónica y fasica de la musculara esquelética que conecta la CV con las extremidades del lado afecto. Un bloqueo que se verá retroalimentado, a su vez, por la falta de impulsos hacia la CV procedentes de las extremidades afectadas. Se trata pues de un circulo vicioso bidireccional: CV-extremidades y extremidades-CV. ¿Como se enfoca esta cuestión en las terapias más utilizadas? Como se ha comentado en los procedimientos actuales los estímulos sobre la MA son unidireccionales, es decir, desde la extremidad/es hacia la CV, con predominio de los movimientos en cadena abierta.
Ejercicios bimanuales
Y funcionan…al menos parcialmente. Parece claro que el movimiento activo de los brazos tiene un efecto sobre la MA de la CV, aunque los trabajos revisados no se ocupen de medir su impacto sobre el tronco (Kim 2017). Entre los que sí lo hicieron destaca el estudio de Lee (2017) con ejercicios bimanuales que tuvo resultados significativos sobre el tronco. En consecuencia realizar movimientos activos diferenciados con los brazos de forma simultánea, tiene un efecto equivalente sobre la MA, con el resultado de un mayor enderezamiento de la CV.
Luego si la actividad diferenciada de los MS provoca un efecto activador en el tronco ¿qué sucedería si se implica en una misma función a las 4 extremidades de forma diferenciada? Lo veremos más adelante, antes es necesario que repasemos algunas cuestiones básicas sobre la MA y su papel en nuestra postura y locomoción.
2. La Musculatura autóctona(MA)
La MA es la verdadera y específica musculatura extensora de la columna. Pertenece a la más antigua musculatura filogenetica, cuya activación no depende de la voluntad. Esta compuesta por diferentes grupos musculares: el tracto lateral (iliocostal y longisimus), el tracto medial (rotadores) y el sistema oblicuo (semiespinosos y Multífidos). Los músculos rotadores y oblicuos de la MA conectan cada una de las vertebras, o bien saltan sobre una o dos de ellas, actuando de esta forma sobre cada uno de los segmentos de la columna. (Sobotta 2006)
El control intersegmental de la columna lo realiza únicamente el tracto medial, cuyo sistema entrelazado constituye una unidad funcional, concebida para una función postural dinámica que se encarga de la extensión de la CV controlando el enderezamiento y por tanto, la movilidad de los cuerpos vertebrales entre sí.
Los músculos del sistema diagonal, ubicados principalmente en el segmento torácico de la CV, juegan un papel decisivo en su extensión axial, aumentando su función postural cuando son traccionados desde la periferia por la fuerza ejercida desde los músculos que trabajan hacia los puntos de apoyo en las extremidades. Esto tendrá un factor capital en la aplicación de la terapia. Este complejo proceso Vojta lo sintetiza así:
«La musculatura autóctona equilibra la columna vertebral, ordenándola intresegmentariamente cuando existen efectos de fuerza extremadamente asimétricos.» El descubrimiento de la motricidad ideal. Pag 205
2.1 Características diferenciales de la MA
La MA tiene una función global sobre la CV debido a que los músculos no están envueltos por fascias, por lo que la activación de un solo músculo irradia hacia la activación automática de todo el paquete muscular. (Vojta-schweizer 2011)
Otra característica diferencial es su inervación realizada por la rama primaria posterior del nervio espinal, toda una división para un reducidisimo numero de músculos: Espinosos, Múltífidos, Rotadores, Longisimus, Intertransversos, e Iliocostal. Considere que la otra división del nervio espinal, la rama primaria ventral, inerva el resto de la musculatura postcraneal (más de 500) ¿Porqué esta gran diferencia? se trata de razones filogeneticas cuando nuestra motricidad como animales acuáticos dependía casi exclusivamente de esta musculatura axial y su activación tenía que preceder temporalmente al resto de la musculatura. Una situación que conservamos actualmente.
La composición histológica de la MA también es especial ya que esta infiltrada de tejido conjuntivo en una alta proporción (25-40%), lo que significa que tiene una gran capacidad de resistencia e información aferente. Debido a esta acumulación de tejido conjuntivo, cuando las vertebras rotan hacia un lado se produce un estiramiento de este tejido (muy limitado en su elasticidad) en los rotadores del lado contrario, acumulando energía para la rotación opuesta con un mínimo de gasto durante la locomoción (Linz 1967). Es este enderezamiento diferenciado de cada uno de los segmentos de la columna el que controla el proceso reciproco de la locomoción hasta las zonas mas distales de las extremidades, y se efectúa de manera automática.
3. La Columna vertebral/MA & extremidades en la filogénesis
La relación entre la MA de la CV y las extremidades se remonta hace 360 millones de años con los primeros anfibios que pasaron su movimiento ondulante acuático al terrestre (Ayala-Conde 2013). Este patrón de locomoción ondulante de los peces es en el que se basa el desplazamiento de los cuadrúpedos, cuya fuerza de propulsión principal es la MA (Edwards 1977). Lo podemos ver en los reptiles y también en los recién nacidos de nuestra especie en una reminiscencia en forma de un reflejo neonatal conocido como Galant. Millones de años de evolución después nuestra locomoción puede llevarnos a tener la falsa impresión que este movimiento ancestral del eje axial ha desaparecido y que toda nuestra locomoción depende del movimiento de nuestras patas traseras.
Tomado de Laufens: Fundamentos biológicos comparativos respecto a la locomoción innata…
Sin embargo, nada mas lejos de la realidad, nuestra MA sigue estando tan interconectada con nuestras extremidades como en nuestros ancestros, aunque ahora ya no vemos las inclinaciones laterales del tronco (salvo en la patología), puesto que en nuestra CV estos movimientos en el plano frontal ahora se realizan en otros planos y con una configuración espacial de la CV adaptada al nuevo tipo de locomoción (Laufens 1991). En síntesis, hemos reconvertido esos patrones mediante la capacidad de nuestra MA de sostenernos extendidos rotando dentro de nuestro eje axial en las secciones torácica y cervical. Un hecho al que Vojta hace referencia del siguiente modo:
«Es seguro que la musculatura autóctona es responsable de la locomoción de los peces y anfibios, provocando un movimiento ondulante que recorre el tronco, y que la marcha vertical del ser humano se basa en el simple perfeccionamiento de dicho movimiento ondulante. Se ha mantenido el principio de organización de la columna vertebral por la musculatura autóctona. Simplemente fue modificado en el transcurso de los largos procesos de la evolución.» El descubrimiento de la motricidad…pag 253
4. La columna vertebral /MA & extremidades en la ontogenesis postural
embrión de 4 semanas
La embriologia nos proporciona importantes pistas sobre como se organiza nuestra compleja arquitectura corporal. Así, a las 4 semanas vemos como aparecen los esbozos de las extremidades, las superiores un poco antes que las inferiores. Son las manos y pies lo primero que germina del tronco y con el crecimiento de las extremidades se van alejando de él. Luego podría decirse que desde un punto de vista embrionario las partes acras están más directamente conectadas con el eje axial que sus prolongaciones (brazos y piernas). Este hecho tendrá su importancia a la hora de explicar determinados fenómenos que observamos en la práctica clínica.
feto a témino
Durante el periodo embrionario, el fetal y en el primer trimestre de vida extrauterina toda la CV permanece en cifosis…el formato estándar de cualquier primate durante toda su vida. A los tres meses de vida, tanto con el patrón del decúbito supino estable, como de forma más evidente con el patrón del apoyo simétrico en codos en decúbito prono, la musculatura axial esta ya completamente desplegada a causa del enderezamiento contra la gravedad, que es cuando el órgano axial es sostenido por las extremidades superiores de apoyo. Entonces los mecanismos de rotación vertebral se hacen necesarios por primera vez, que en cifosis se encuentran muy limitados. La MA mantendrá durante toda la ontogenesis postural una relación sinérgica con el resto de la musculatura esquelética (Vojta-Peters 1992)
Postura a los 3 meses
A partir de este primer trimestre la MA continuará diferenciándose más y más en sincronía con los nuevos patrones que van madurando hasta alcanzar la marcha bípeda. El enderezamiento intersegmentario de la columna vertebral comienza en la zona craneal e implica a todos sus segmentos. Que se produzca la diferenciación funcional de los músculos que organizan la actividad dinámica de cada segmento de la musculatura vertebral depende la diferenciación del resto de la musculatura esquelética, incluyendo la motricidad fina.
No podemos olvidar que la calidad de la postura y la motricidad están en relación con la calidad en el control dinámico de la columna vertebral. Vojta lo resume de esta forma: «La diferenciación de la musculatura autóctona de la columna, una musculatura antiquísima de los vertebrados, es la base en que se apoya la diferenciación de la actividad muscular de todo el cuerpo.» El Principio Vojta
5. Un enfoque diferente: La Locomoción refleja de Vojta y su efecto sobre la MA
Con la aplicación terapéutica de la LR tenemos la posibilidad de activar directamente y de forma refleja la MA, llevándola a extender axialmente la CV con la finalidad de facilitar su diferenciación dentro de un patrón global innato de tipo locomotor como es la Reptación refleja. (Vojta 1991)
5.1Activación refleja de la MA mediante las Zonas de la LR.
Vojta clasifico las zonas de activación (descubiertas de forma empírica) en dos categorías. A las encontradas en las extremidades las denominó zonas principales, y a las situadas en el tronco, secundarias (Vojta-Peters 1992). Una distinción formal sin valor en la terapia, ya que no implica predominancia de unas sobre otras, aunque Vojta precisaba que: «la experiencia clínica ha demostrado que, en el recién nacido sano, todo el complejo de coordinación aparece más deprisa y de forma más completa desde las zonas principales que desde las secundarias.»
¿Qué puede significar? parece confirmar que la generación refleja de puntos de apoyo en las extremidades tiene un potente efecto activador de la MA anticipando una respuesta postural en el eje axial que no estará disponible en la motricidad espontánea del bebé hasta mediados del 2º trimestre, y cuyos patrones parciales abarcan desde el primer trimestre hasta la marcha bípeda. (Vojta-Peters 1992)
Esto ha sido observado niños y adultos sanos, pero si tenemos presente que en el daño cerebral se ve seriamente comprometida la función enderezadora de la CV por parte de la MA, esta situación tendrá su correspondiente impacto sobre las extremidades, cuya afectación, a su vez, suprimirá los impulsos hacia la CV.
¿Como salir de este circulo vicioso? ya hemos visto cómo las terapias más eficaces en neurorehabilitación abordan esta cuestión, como la terapia bimanual que activa el enderezamiento axial mediante los impulsos generados por la actividad diferenciada de ambos MS. La LR lo enfoca de forma diferente.
La locomoción refleja de Vojta implica a las 4 extremidades generando impulsos simultáneamente y de forma diferenciada desde cada una de ellas hacia la CV. Esta diferenciación corresponde a que cada extremidad, al estar integrada en el contexto de un patrón de locomoción cuadrúpeda, (como es la Reptación refleja) se encuentran en distintos momentos del ciclo del paso: Flexión, relajación, apoyo e impulso (Grillner 1981). La CV recibe estos impulsos ya extendida de forma refleja (por el tracto medial de la MA), condición necesaria para ejecutar los movimientos de rotación recíprocos que proporcionan el soporte postural a los movimientos de las extremidades (Vojta-Peters 1991). Esta sincronía bidireccional entre MA y extremidades es la que se produce de forma natural en las locomociones humanas y durante toas las actividades fucionales de los MS.
Veamos como se produce esta activación de la MA de forma detallada en el patrón de la Reptación Refleja.
5.1 A) Las zonas secundarias y la MA
En el tronco Vojta y colaboradores encontraron 5 zonas simétricas (cada zona tiene su equivalente en el otro lado): dos en la escapula: acromion (nucal) y borde medial (facial); dos en la pelvis: EIAS (facial) y Zona glútea (nucal); y una en el tórax: la zona del tronco ya mencionada (nucal). Todos activan el mismo patrón con acceso directo a la MA:
Combinación acromión, glúteo
La zonaacromioncon su vector medial ofrece un input de aproximación de la escapula hacia la CV, activando los addutores en sentido medial, es decir, con punto fijo en la CV para lo cual la MA es requerida para mantenerse extendida en su eje de forma dinámica para permitir la rotación.
La zona del tronco con su strecht directo sobre las articulaciones costotransversas, provocando la distensión/activación de la MA.
La zonaborde medial de la escapula que con su vector lateral hacia el codo facial de apoyo provoca la activación refleja de los addutores en sentido lateral, es decir, hacia el apoyo en el codo facial, provocando con ello la rotación de las espinosas dorsales. Esta tracción tiene un potente efecto activador de la MA que debe mantenerse en su eje en la zona dorsal para facilitar esta rotación.
La zona Espina Iliaca Antero Superior (EIAS) activa la MAtravés del strecht sobre la inserción de los abdominales (transverso y oblicuos) en esta zona y su conexión directa con la CV lumbar mediante su fascia de inserción en los procesos transversos lumbares. La activación de la MA en la región lumbar por la zona EIAS es especialmente importante por la fuerte asimetría de las fuerzas que operan en esta sección de la CV, bien para provocar hiperlordosis, cifosis o bien una incurvación excesiva en el lado facial. Por tanto, esta zona no solo tiene su influencia en la musculatura ventral, también, y de manera decisiva, sobre la MA.
Combinación cabeza con zona glutea
La zona glútea se relaciona directamente con la MA a través de la activación del giro del iliaco hacia dorsal, caudal y lateral, actuando sobre distintos niveles: en la articulación sacro iliaca; sobre la inserción en el sacro del tracto lateral de la MA y la fascia toraco-lumbar; y sobre el cuadrado lumbar que se relaciona directamente con la MA.
5.1 B) Las zonas principalesy la MA
La relación de las zonas principales con la MA, esta en función del rol que esa extremidad ocupa en el patrón de locomoción global, que pueden ser de dos tipos: Tónicas o fásicas. Todas las zonas tienen en común un vector medial (hacia la CV) con la ambigüedad en la estiloides radial por la posición flexionada del codo.
Combinación talon con escapula facial
En el caso de las zonas tónicas o de apoyo (talón nucal y codo facial) la MA es activada como consecuencia de las fuerzas que traccionan del tronco, y por tanto de la CV hacia las extremidades de apoyo. Se trata de una respuesta de enderezamiento axial que se encarga de sostener la CV en su eje axial para hacer más eficientes las fuerzas que proyectan la CV y a todo el cuerpo en el sentido de la locomoción. Las desviaciones que se produzcan fuera de este eje axial reflejarán los déficits de la MA observándose sus compensaciones más evidentes en las extremidades durante esta fase de apoyo.
Cuando la zona talón se constituye como punto fijo, su efecto sobre la MA se relaciona más con la región lumbar del lado nucal, ejercida por el psoas y cuadrado lumbar de este lado, cuyos vectores de contracción se dirigen hacia sus inserciones distales, provocando con ello fuerzas generadoras de excurvación (esta zona no puede rotar). Estas fuerzas serán contrarrestadas por el strecht que esta distensión produce sobre la MA, provocando su activación enderezadora de la CV.
La zona del codo facial actúa de forma similar a la del talón sobre la MA, esta vez en la región torácica de la CV. Esta región, al contrario de la columna lumbar, responde rotando sus espinosas hacia el punto de apoyo en el codo facial. Algo que solo es posible si la CV es mantenida en extensión por la MA, extensión provocada a su vez por esta acción rotadora, junto con la propia activación de la MA generada desde la CV.
En las zonas fásicas la estiloides del radio (nucal) tiene su efecto sobre la CV torácica de forma indirecta a través de la activación de los rotadores externos, abd y flexores del hombro. Este movimiento de avance de la extremidad nucal es el que provoca la rotación de los cuerpos vertebrales torácicos hacia este lado. De nuevo, esta rotación implica la activación de la MA como requisito para que dicha rotación tenga lugar dentro del eje axial, junto con la propia activación enderezadora generada desde la CV.
La zona del cóndilo medial del fémur tiene su efecto sobre la MA a través de la activación de los rotadores externos, abductores y flexores de la cadera de la cadera facial, y también de forma indirecta a los abdominales. Sin embargo, el movimiento de avance de esta pierna necesita que la MA este previamente activada. A ello contribuye la fascia toracolumbar, dorsal ancho y cuadrado lumbar del lado facial que al tener su vector de tracción hacia el codo de apoyo facial actúan controlando el grado de incurvación lumbar de la CV durante la flexión de la pierna. Esta enorme concentración de fuerzas con vectores opuestos sobre esta región exigen la máxima activación diferenciada de la MA en esta zona.
5.2 Consideraciones prácticas para la activación de la MA desde las zonas en la LR
Combinación triple: talón, glútea y codo.
La cabeza durante la realización de la terapia, es considerada una proyección de la CV, de hecho, Vojta la definía como el «órgano efector» o de resultado de la activación refleja, ya que expresa fielmente con su alineación (o desviación) en el eje axial, y su movimiento de rotación, la calidad y cantidad de la activación de la MA (que no podemos ver), así como el momento del ciclo del paso en el que se encuentra el patrón global.
Por la misma razón, el proceso también opera a la inversa, es decir, guiar la posición de la cabeza en la terapia tiene un efecto directo sobre la activación de la MA, e influye sobre el momento del ciclo del paso sobre el que queremos incidir.
Por tanto, controlar la cabeza antes de activar el patrón global, posicionándola lo más centrada en el eje axial, y guiándola durante la activación para mantenerla dentro de su eje, resistiendo o facilitando su rotación (según convenga) hace de la cabeza una de las «zonas» más importantes en la activación de la MA. Considere que en el patrón de la RR, la cabeza inicia el giro hacia el lado nucal cuando se constituye el codo facial como punto de apoyo. La resistencia a este giro incrementará la activación del punto de apoyo intensificando la contracción de la MA para mantenerse en el eje axial al ser giradas las espinosas del segmento dorsal hacia el punto fijo: el codo apoyado.
El control de la cabeza es clave en el proceso de activación de la MA de la Columna
La zona del tronco no es una zona más, ya que tiene acceso directo a la activación de la MA. Se trata de una combinación de estimulo perióstico con stretch muscular de carácter propioceptivo. Aplicado de forma aislada tiene un efecto enderezador de la CV, más intenso en el segmento dorsal. En la RR se utiliza en el lado nucal e influye, además de en la extensión de la CV dorsal, en la add de la escapula nucal, rotación externa del hombro nucal y en su movimiento de avance. Una acción que se ve acelerada en combinación con la zona acromion, también nucal.
En el decúbito lateral, perteneciente al volteo reflejo, la zona del tronco aplicada en el lado de arriba ejerce idéntico efecto sobre la CV dorsal, y similar sobre el brazo de arriba en combinación con otras zonas, normalmente la EIAS del lado de arriba.
Combinación zona del tronco + acromión
¿Qué pasa si se aplica esta zona del tronco en el lado facial de la RR? la experiencia clínica muestra que se activa la MA de este lado facial de forma diferenciada para sostener la CV en su eje mientras sus espinosas son rotadas en la dirección de este brazo facial, donde se produce el apoyo en el epicondilo medial del húmero. Esta es probablemente la explicación de que esta zona se aplique en el lado nucal en lugar de hacerlo en el facial, puesto que en el lado facial se encuentra el punto de apoyo donde la musculatura adductora de la escapula (trapecio y romboides) será la responsable de hacer girar las espinosas hacia este apoyo, activando con ello a la MA que tendrá que sostener la CV dentro de su eje evitando que se produzca una excurvación.
Sin embargo, en caso de hemiparesia habría que considerar que también es necesario estimular la zona del tronco también en el lado facial, puesto que no será tan sencillo lograr el sostenimiento de la CV dentro de su eje longitudinal por la sola acción procedente del apoyo en el MS afecto, también muy difícil de conseguir. La dirección del estímulo de esta zona del tronco en el lado facial se ha mostrado más efectiva, en casos clínicos, hacia el punto de apoyo en lugar de hacerlo hacia la CV (como cuando se estimula siendo nucal)
En cualquier caso se trata de buscar una forma de activar la MA de forma diferenciada al servicio de la función.
«La terapia Vojta puede definirse como medida básica y selectiva para la diferenciación de la musculatura autóctona» El descubrimiento de la motricidad ideal, pag 254
6. Consideraciones finales
En la patología neurológica la rotación segmental de las vertebras entre sí esta muy limitada en la región torácica (relacionada con las funciones del MS), en tanto en cuanto esta afectada la capacidad de extenderse axialmente, una función esencial de la MA. En consecuencia, el desarrollo motor normal, así como su mantenimiento en la edad adulta depende de la actividad eficiente de la MA.
Las metodologías que abordan las secuelas sensorio-motoras del MS a través del movimiento activo, sea cual sea el procedimiento, tienen su reflejo más o menos efectivo sobre el tronco. Se trata de impulsos unidireccionales hacia la CV que estimulan la contracción de la MA en su segmento torácico, y que en el mejor de los casos proceden de 2 extremidades al tiempo.
Con la aplicación de la LR, a diferencia de las terapias comentadas, se ofrece impulsos altamente diferenciados desde las 4 extremidades hacia la CV, y desde la CV hacia las extremidades, (estímulos bidireccionales) de forma refleja con efectos muy positivos en la función de los MS y en la marcha (Laufens 1996, Backtroem 2000, Husarova 2005, Perales 2009, 2011, 2013, Epple 2018). Sin embargo, la evidencia científica aún no es suficiente como para afirmar su superioridad en relación a otros procedimientos.
«El que los mecanismos de rotación de la CV [dependientes de la MA] puedan actuar sobre sobre una columna extendida es la preparación necesaria para que puada darse una locomoción bípeda coordinada.» El principio Vojta, pag 144
Bibliografia
Kwakkel G, Kollen BJ, van der Grond J, Prevo AJ. Probability of regaining dexterity in the flaccid upper limb: impact of severity of paresis and time since onset in acute stroke. Stroke. 2003;34(9):2181-2186. 2
Dixit S, Tedla JS. Effectiveness of robotics in improving upper extremity functions among people with neurological dysfunction: a systematic review. Int J Neurosci. 2019 Apr;129(4):369-383.
Sale P, Mazzoleni S, Lombardi V, Galafate D, Massimiani MP, Posteraro F, Damiani C, Franceschini M. Recovery of hand function with robot-assisted therapy in acute stroke patients: a randomized-controlled trial.Int J Rehabil Res. 2014 Sep;37(3):236-42.
Aprile I, Germanotta M, Cruciani A, Loreti S, Pecchioli C, Cecchi F, Montesano A, Galeri S, Diverio M, Falsini C, Speranza G, Langone E, Papadopoulou D, Padua L, Carrozza MC. Limb Robotic Rehabilitation After Stroke. A Multicenter, Randomized Clinical. Journal of Neurologic Physical Therapy: January 2020 – Volume 44 – Issue 1 – p 3–14
Veerbeek JM, Langbroek-Amersfoort AC, van Wegen EE, Meskers CG, Kwakkel G.Effects of Robot-Assisted Therapy for the Upper Limb After Stroke. Neurorehabil Neural Repair. 2017 Feb;31(2):107-121.
Laffont I, Froger J, Jourdan C, Bakhti K, van Dokkum LEH, Gouaich A , Bonnin HY , Armingaud P, Jaussent A , Picot MC , Le Bars E , Dupeyron A, Arquizan C, Gelis A , Mottet D. Rehabilitation of the upper arm early after stroke: video games versus conventional rehabilitation. A randomized controlled trial. Ann Phys Rehabil Med. 2019 9 de diciembre. Pii: S1877-0657 (19) 30188-5.
Brunner I, Skouen JS, Hofstad H, Aßmus J, Becker F, Sanders AM, Pallesen H, Qvist Kristensen L, Michielsen M, Thijs L, Verheyden G. Virtual Reality Training for Upper Extremity in Subacute Stroke (VIRTUES): A multicenter RCT. Neurology. 2017 Dec 12;89(24):2413-2421.
Chang KV, Wu WT, Huang KC, Han DS. Segmental body composition transitions in stroke patients: Trunks are different from extremities and strokes are as important as hemiparesis Clin Nutr. 2019 Aug 31. pii: S0261-5614(19)33036-5.
Alhwoaimel N, Turk R, Warner M, Verheyden G, Thijs L, Wee SK, Hughes AM. Do trunk exercises improve trunk and upper extremity performance, post stroke? A systematic review and meta-analysis. NeuroRehabilitation. 2018;43(4):395-412.
Thrane G, Alt Murphy M, Sunnerhagen KS. Recovery of kinematic arm function in well-performing people with subacute stroke: a longitudinal cohort study. J Neuroeng Rehabil. 2018 Jul 18;15(1):67.
Razmus M. Body representation in patients after vascular brain injuries. Cogn Process. 2017 Nov;18(4):359-373.
Fernandez-Gomez E, Sanchez-Cabeza A. Motor imagery: a systematic review of its effectiveness in the rehabilitation of the upper limb following a stroke. Rev Neurol. 2018 Mar 1;66(5):137-146.
Yadav RK, Sharma R, Borah D, Kothari SY. Efficacy of Modified Constraint Induced Movement Therapy in the Treatment of Hemiparetic Upper Limb in Stroke Patients: A Randomized Controlled Trial. J Clin Diagn Res. 2016 Nov;10(11):YC01-YC05.
Etoom M1, Hawamdeh M, Hawamdeh Z, Alwardat M, Giordani L, Bacciu S, Scarpini C, Foti C. Constraint-induced movement therapy as a rehabilitation intervention for upper extremity in stroke patients: systematic review and meta-analysis. Int J Rehabil Res. 2016 Sep;39(3):197-210.
Lee DH1, Park SH1, Han JW1. Effect of bilateral upper extremity exercise on trunk performance in patients with stroke. J Phys Ther Sci. 2017 Apr;29(4):625-628.
Kim J1, Yi J2, Song CH1. Kinematic analysis of head, trunk, and pelvic motion during mirror therapy for stroke patients. J Phys Ther Sci. 2017 Oct;29(10):1793-1799.
Vaclav Vojta, Annegret Peters. El principio Vojta. 1992. Springer Verlag-Iberica.
Sobotta, J. (2006). Atlas de anatomia humana (Vol. 2). Ed. Médica Panamericana.
Vojta V, Schweizer E. El descubrimiento de la motricidad ideal. Ediciones Morata 2011.
Edwards J.L, 1977. The evolution of terrestrial locomotion. In: Hecht, M.K, Goody, P.C, and B.M. Hecht(Eds): Major patterns in vertebrate evolution, 533-577. New York.
Grillner S (1981) Control of locomotion in bipeds, tetrapods and fish. In: Brookhart JMaMVB (ed) Handbook of physiology. The nervous system. Motor control. American Physilogy Society, New York, pp 1179–1236.
G. Laufens, S. Seitz, G. Staenicke, Fachbereich. Fundamentos biológicos comparativos respecto a la locomoción innata, en particular respecto a la “reptación refleja” según Vojta. Fisioterapia 43. Jg (5/1991), Pag. 448-156.
Laufens G, Reimann G., Poltz W., Schmiegelt F.: Behandlunserfolge und Bedingungen der VojtaPhysio-Therapie bei MS-Patienten. (Exito en el tratamiento y condiciones de la fisioterapia Vojta en pacientes con esclerosis multiple) Krankengymnastik 4, 1996. 518-532.
Backstrom B. Vojta self-training: experiences of six neurologically impaired people. Physiotherapy. 2000;86(11):567-74.
Husárová, R. The vojta approach in adult patients. Rehabilitacia. 2005;42(3).
Perales L, Pérez AM, Atim MA, Varela E. Efecto de la terapia Vojta en la rehabilitación de la marcha en dos pacientes adultos con daño cerebral adquirido en fase tardía. Fisioterapia. 2009;31:151-62.Pavlu D, Véle F, HavlíˇckováL . Electromyography and kinesiological analysis of Vojta´s therapeutical principle. Rehab Fyzil. 2000;7:34-9.
Perales-López, L. El pisiforme, una nueva zona de desencadenamiento segmental de los contenidos cinesiológicos de la mano en la terapia Vojta. Aplicaciones en rehabilitación. Fisioterapia, Vol 35, (5), P 189-196.
Perales-López L, Fernández-Acenero M.J. ¿Es transferible la terapia de locomoción refleja a una plataforma de teleneurorrehabilitación en el tratamiento del paciente adulto? Rehabilitación . 2013;47(4):205-212.
Epple C, Maurer-Burkhard B, Lichti MC and Steiner T. Vojta Therapy in Patients with Acute Stroke – A New Approach in Stroke Rehabilitation. Phys Med Rehabil Int. 2018; 5(1): 1140.